Vous aimerez peut-être aussi
- Comprendre Toute La FinanceDocument26 pagesComprendre Toute La Financeمرح و فرفشة50% (2)
- Les Chakras Et Le Corps Éthérique (Coquet Michel)Document217 pagesLes Chakras Et Le Corps Éthérique (Coquet Michel)Teddy Ravatsy100% (1)
- QE de Physiologie EndocrinoDocument12 pagesQE de Physiologie EndocrinoN.Pas encore d'évaluation
- Exploration de L'axe GonadotropeDocument113 pagesExploration de L'axe Gonadotropeyolene mumbaPas encore d'évaluation
- 1 As - Projet 1 Entier (5 Séquences)Document74 pages1 As - Projet 1 Entier (5 Séquences)HacerMellal83% (36)
- Le Système EndocrinienDocument23 pagesLe Système Endocrinienchadi ouechtatiPas encore d'évaluation
- Glande EndocrineDocument30 pagesGlande EndocrineMACON824Pas encore d'évaluation
- Cours 6Document27 pagesCours 6BouabidiPas encore d'évaluation
- Le Cycle Sexuel Chez La Femme TCDocument9 pagesLe Cycle Sexuel Chez La Femme TCSarah DiomandePas encore d'évaluation
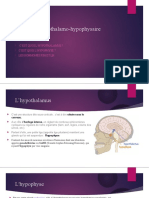
- Physiologie Du Complexe Hypothalamo-HypophysaireDocument5 pagesPhysiologie Du Complexe Hypothalamo-HypophysaireZied Bouabene100% (1)
- L'axe Hypothalamo-HypophysaireDocument11 pagesL'axe Hypothalamo-Hypophysairegeorge.guergourPas encore d'évaluation
- UE8 TC Physio Cours 2 - Régulation Endocrinienne Tutorat 2013-2014Document10 pagesUE8 TC Physio Cours 2 - Régulation Endocrinienne Tutorat 2013-2014Ali EL Kihel100% (1)
- Exploration de L Axe GonadotropeDocument14 pagesExploration de L Axe GonadotropeDjouhaina BeggahPas encore d'évaluation
- Rapport de Stage Realisation D Un ERPDocument51 pagesRapport de Stage Realisation D Un ERPmohamed benaichPas encore d'évaluation
- Axe Hypothalamo-Hypophysaire (Resume)Document7 pagesAxe Hypothalamo-Hypophysaire (Resume)Maroua BenchoufiPas encore d'évaluation
- Grand Oral SesDocument2 pagesGrand Oral Sesapi-657069168Pas encore d'évaluation
- A.Saudreau Les Degrees de Vie Spirituel 1 PDFDocument596 pagesA.Saudreau Les Degrees de Vie Spirituel 1 PDFpbelo1Pas encore d'évaluation
- Correction Du Devoir IntroductionDocument5 pagesCorrection Du Devoir IntroductionMme et Mr LafonPas encore d'évaluation
- 02-PHYSIOLOGE DE L'AXE HYPOTHALAMO HYPOPHYSAIRE Poly-1Document3 pages02-PHYSIOLOGE DE L'AXE HYPOTHALAMO HYPOPHYSAIRE Poly-1alimkettouchePas encore d'évaluation
- PHYSIOLOGE DE L'AXE HYPOTHALAMO HYPOPHYSAIRE PolyDocument4 pagesPHYSIOLOGE DE L'AXE HYPOTHALAMO HYPOPHYSAIRE PolyFatima zahra NaciriPas encore d'évaluation
- Régulation Physiologique Du Complexe Hypothalamo - HypophysaireDocument15 pagesRégulation Physiologique Du Complexe Hypothalamo - HypophysaireRyanPas encore d'évaluation
- Le Systeme EndocrinienDocument13 pagesLe Systeme EndocrinienAlexiPas encore d'évaluation
- Axe Hypothalamo HypophysaireDocument4 pagesAxe Hypothalamo HypophysaireAbderahmene GhodbanePas encore d'évaluation
- 3-Cours Endocrinologie Et Régulation Des Systèmes Chapitre 2 M1 TOXICOLOGIE 2020-2021 BOUBEKRIDocument32 pages3-Cours Endocrinologie Et Régulation Des Systèmes Chapitre 2 M1 TOXICOLOGIE 2020-2021 BOUBEKRIBERDJOUH YoucefPas encore d'évaluation
- MNE CM3bis Physiologie Axe Hypothalamo HypophysaireDocument13 pagesMNE CM3bis Physiologie Axe Hypothalamo HypophysaireSoheib RahrahPas encore d'évaluation
- Cours - 3 Les Régulation Hormonale de La MétabolismeDocument9 pagesCours - 3 Les Régulation Hormonale de La Métabolismemiss anthropocenePas encore d'évaluation
- GYNECO Prof MBUYAMBA - 090159Document189 pagesGYNECO Prof MBUYAMBA - 090159reneddymutombo22Pas encore d'évaluation
- 2-Complexe Hypothalamo-Hypophysaire PDFDocument26 pages2-Complexe Hypothalamo-Hypophysaire PDFthierno amadou bahPas encore d'évaluation
- Physiologie de L'Hypophyse: MASTER 1 Médecine 2023Document61 pagesPhysiologie de L'Hypophyse: MASTER 1 Médecine 2023Lorlyna MandzeyiPas encore d'évaluation
- 62 Coursts 08Document4 pages62 Coursts 08Nicoleta ChivuPas encore d'évaluation
- Le Systeme Endocrinien WDocument9 pagesLe Systeme Endocrinien WsafemindPas encore d'évaluation
- Physio Endocrino23-01glandes EndocrinesDocument4 pagesPhysio Endocrino23-01glandes EndocrinesVictorin FANDREBOPas encore d'évaluation
- Ahh M2decineDocument22 pagesAhh M2decinemalakzerizer2005Pas encore d'évaluation
- Notions D'endocrinoDocument119 pagesNotions D'endocrinoyaya camaraPas encore d'évaluation
- La Reproduction Chez Les Mammiferes Resume Lecon2 Module 2Document6 pagesLa Reproduction Chez Les Mammiferes Resume Lecon2 Module 2Antoine S.Pas encore d'évaluation
- Axe HypothalmoDocument43 pagesAxe HypothalmoDoc BelhadjPas encore d'évaluation
- Hypophys e 4Document32 pagesHypophys e 4trx100% (1)
- TP 2 1 Biologie AnimaleDocument7 pagesTP 2 1 Biologie Animaleazertyiashmy1234Pas encore d'évaluation
- Cours Final SomatostatineDocument4 pagesCours Final Somatostatineemawatsn122Pas encore d'évaluation
- 2 Relations Hypothalamus Hypophyse BDocument27 pages2 Relations Hypothalamus Hypophyse BOXGAMINGXOPas encore d'évaluation
- Cours Endocrinologie de La ReproductionDocument49 pagesCours Endocrinologie de La ReproductionLukusa MauricePas encore d'évaluation
- Chapitre V MineDocument19 pagesChapitre V MineMer IemPas encore d'évaluation
- Principes Des Systemes de Controle HormonalDocument2 pagesPrincipes Des Systemes de Controle Hormonalfafoufa133Pas encore d'évaluation
- Hypothalamo HypophysaireDocument8 pagesHypothalamo HypophysaireMaktabatiLB PlannersPas encore d'évaluation
- Les Neurohormones Chez HommeDocument17 pagesLes Neurohormones Chez HommeNisrine 2112Pas encore d'évaluation
- Cours 3Document16 pagesCours 3azizhseck2Pas encore d'évaluation
- Le Système Hypothalamo-Hypophysaire: DR Souleymane THIAM Assistant Biochimie L2S3Document54 pagesLe Système Hypothalamo-Hypophysaire: DR Souleymane THIAM Assistant Biochimie L2S3s22g9ndkwjPas encore d'évaluation
- PDF Document 3Document3 pagesPDF Document 3Fatou Bintou RassoulPas encore d'évaluation
- Axe HPT-HPPDocument36 pagesAxe HPT-HPPRezagui ImadPas encore d'évaluation
- 5-La Corticosurrénale (CS)Document32 pages5-La Corticosurrénale (CS)DëLux OwOPas encore d'évaluation
- 2014 7 Le Systeme EndocrinienDocument15 pages2014 7 Le Systeme EndocrinienSamira MBasskarPas encore d'évaluation
- Endocrinologie IDocument88 pagesEndocrinologie Iritajbeily1611Pas encore d'évaluation
- Révisions 2.2Document16 pagesRévisions 2.2lenapaquiePas encore d'évaluation
- 2-Axehthp M1bse EtudiantsDocument50 pages2-Axehthp M1bse EtudiantsHq sishPas encore d'évaluation
- Document de L'hypothalamus, Fiche MedecineDocument1 pageDocument de L'hypothalamus, Fiche Medecine00mohamedjbeli00Pas encore d'évaluation
- T3A2 Sexualite Et ProcreationDocument34 pagesT3A2 Sexualite Et ProcreationBechirMoussiPas encore d'évaluation
- Relation Hypothalamo-HypophysaireDocument8 pagesRelation Hypothalamo-HypophysaireAsla AsmaPas encore d'évaluation
- Endocrinologie Fœto-PlacentaireDocument21 pagesEndocrinologie Fœto-PlacentaireNour ALPas encore d'évaluation
- Fichier Produit 2218Document5 pagesFichier Produit 2218Alycia SafaPas encore d'évaluation
- 5.0 Endocrinologie NaassilaDocument62 pages5.0 Endocrinologie NaassilaMohamed AraouchiPas encore d'évaluation
- T3A2 Acti2 FCTT OvairesDocument3 pagesT3A2 Acti2 FCTT Ovairesbianca fPas encore d'évaluation
- Retenir Seq 5 Part 3Document6 pagesRetenir Seq 5 Part 3Julia NicolettiPas encore d'évaluation
- Physiologie Endocrinienne m1023Document71 pagesPhysiologie Endocrinienne m1023liliahadjali57Pas encore d'évaluation
- 9.fonctions Gonadiques FémininesDocument51 pages9.fonctions Gonadiques FémininesAmmar ChaouiPas encore d'évaluation
- Axe Hypothalamo-HypophysaireDocument25 pagesAxe Hypothalamo-HypophysaireAnfelDiivaPas encore d'évaluation
- TD1 Regulation SABRINA.2019Document5 pagesTD1 Regulation SABRINA.2019axelPas encore d'évaluation
- Hormones HypothalamiquesDocument14 pagesHormones Hypothalamiquesdjamila benaissaPas encore d'évaluation
- Activité - Les Pièces de La MaisonDocument4 pagesActivité - Les Pièces de La MaisonlakhanicherishalPas encore d'évaluation
- Demande de StageDocument1 pageDemande de StageIbrahima SkPas encore d'évaluation
- Evaluation Diagnostique - 1 BACSDocument2 pagesEvaluation Diagnostique - 1 BACSmeryemPas encore d'évaluation
- Etat Et JusticeDocument12 pagesEtat Et Justicenoa.blain49Pas encore d'évaluation
- Bounty Maison LightDocument2 pagesBounty Maison LightArribardPas encore d'évaluation
- (Brico) Tridome - 19 Poser Une FenêtreDocument2 pages(Brico) Tridome - 19 Poser Une FenêtreMadidj_2014Pas encore d'évaluation
- BOMP 1525 Du Mardi 13 Août 2019Document84 pagesBOMP 1525 Du Mardi 13 Août 2019Moussa Karim AlioPas encore d'évaluation
- EcoChap2 Défaillances Etat Marché Découvrir Cours Hamza MustafaDocument3 pagesEcoChap2 Défaillances Etat Marché Découvrir Cours Hamza Mustafahamza attariPas encore d'évaluation
- FAQ OBD-Honda FRDocument3 pagesFAQ OBD-Honda FRCarlos SoaresPas encore d'évaluation
- L'ego-Histoire en PerspectiveDocument11 pagesL'ego-Histoire en PerspectiveNicoleta AldeaPas encore d'évaluation
- Art Du MontageDocument18 pagesArt Du MontageAndré FélixPas encore d'évaluation
- Offre de ServiceDocument6 pagesOffre de Services.elgasmiPas encore d'évaluation
- Bertrand, Alexandre Louis Joseph (1820-1902) - La Religion Des Gaulois Les Druides Et La Druidisme (E. Leroux, 1897)Document394 pagesBertrand, Alexandre Louis Joseph (1820-1902) - La Religion Des Gaulois Les Druides Et La Druidisme (E. Leroux, 1897)highrangePas encore d'évaluation
- Correction Projets PowerPointDocument16 pagesCorrection Projets PowerPointrakez bassemPas encore d'évaluation
- ArticleDocument18 pagesArticleAbderrahim ElPas encore d'évaluation
- Cps Type Des Marches de Travaux 2015Document29 pagesCps Type Des Marches de Travaux 2015azoughPas encore d'évaluation
- 9.douleur ThoraciqueDocument22 pages9.douleur ThoraciqueFerhani OuaPas encore d'évaluation
- Orca Share Media1663662719481 6977907198972110101Document43 pagesOrca Share Media1663662719481 6977907198972110101Bilel LoussifPas encore d'évaluation
- Fiches TD Micro IIIDocument4 pagesFiches TD Micro IIIABRAHAM NENE100% (1)
- QCM Séisme À ImprimerDocument18 pagesQCM Séisme À ImprimerDieudonné AdokoPas encore d'évaluation
- Daressy Statues I 1906Document427 pagesDaressy Statues I 1906Eugenia MuñozPas encore d'évaluation
- Les Imperfections de L Oeil 2Document5 pagesLes Imperfections de L Oeil 2s18db911Pas encore d'évaluation
- RCAD Acier Manuel FR 50Document284 pagesRCAD Acier Manuel FR 50Nanescu LilianaPas encore d'évaluation