Vous aimerez peut-être aussi
- OsmoseDocument14 pagesOsmoseValéry Epee Ebongue100% (1)
- Osmose InverseDocument10 pagesOsmose InverseHassan FekiPas encore d'évaluation
- Chapitre V Osmose InverseDocument11 pagesChapitre V Osmose InverseHassanPas encore d'évaluation
- Osmose InverseDocument50 pagesOsmose InverseIkram Ziri50% (2)
- Rhéologie 110524Document11 pagesRhéologie 110524alnutsPas encore d'évaluation
- Exercices Tensioactifsemulsions MoussesDocument3 pagesExercices Tensioactifsemulsions MoussesYassine RakchoPas encore d'évaluation
- Les Utils Théoriques de La Mécanique Des FluidesDocument24 pagesLes Utils Théoriques de La Mécanique Des FluideslorenzodecocoabdouPas encore d'évaluation
- Chap 5 DessalementDocument106 pagesChap 5 DessalementAbdou DidosPas encore d'évaluation
- Osmose InverseDocument12 pagesOsmose InverseSlim Kanoun100% (1)
- OsmomètrieDocument16 pagesOsmomètrieredwanePas encore d'évaluation
- Le Milieu InterieurDocument30 pagesLe Milieu InterieurMedMan6100% (9)
- ACFrOgBKzf1385QSRlwPvvXSBbVxzjPV99GpWv5bmptzYDYFGVQwxJPMxtiBXp8CqfemclZHdKw7SSQVpCUn025sSybqmMEkXKyjWGW8Vh9JbGjM4P3xH2psWYdarcZxzBaVSingKAsFB3TR7j U PDFDocument10 pagesACFrOgBKzf1385QSRlwPvvXSBbVxzjPV99GpWv5bmptzYDYFGVQwxJPMxtiBXp8CqfemclZHdKw7SSQVpCUn025sSybqmMEkXKyjWGW8Vh9JbGjM4P3xH2psWYdarcZxzBaVSingKAsFB3TR7j U PDFBouchra BkhtmPas encore d'évaluation
- Traitement - Des - Accidents - Dus - Lextravasa20210422 25976 Hd8om4 With Cover Page v2 PDFDocument18 pagesTraitement - Des - Accidents - Dus - Lextravasa20210422 25976 Hd8om4 With Cover Page v2 PDFMohamed HamilaPas encore d'évaluation
- Bioph2an16-Rheologie CherefDocument21 pagesBioph2an16-Rheologie Cherefishaq AllalPas encore d'évaluation
- Cours DSVT Terminal CDocument33 pagesCours DSVT Terminal CMichael JamesPas encore d'évaluation
- Procédés Membranaires Pour Le Traitement de L'eauDocument44 pagesProcédés Membranaires Pour Le Traitement de L'eauSara Qrm100% (1)
- Chapitre 06 Osmose de DiffusionDocument6 pagesChapitre 06 Osmose de DiffusionsoufianePas encore d'évaluation
- Tpe Mariecur804eDocument29 pagesTpe Mariecur804eHAJARPas encore d'évaluation
- Mecanismes D Absorption de L Eau Et Des Sels Mineraux Chez Les Plantes Cours 1Document25 pagesMecanismes D Absorption de L Eau Et Des Sels Mineraux Chez Les Plantes Cours 1Mohamed ACHAHBAR100% (3)
- Cours3 18oct15Document24 pagesCours3 18oct15yas minePas encore d'évaluation
- Techniques Des FluidesDocument14 pagesTechniques Des FluidesNdendenguiPas encore d'évaluation
- ArgileDocument14 pagesArgilePeter TahaPas encore d'évaluation
- Membranes OIDocument14 pagesMembranes OIOliver FrerePas encore d'évaluation
- Meca-Flu3 Mécanique Des FluidesDocument22 pagesMeca-Flu3 Mécanique Des FluidesAhmad CisséPas encore d'évaluation
- Extraitenseignementscientifique 1 EreDocument8 pagesExtraitenseignementscientifique 1 EreKhadimoul Moustapha NiangPas encore d'évaluation
- Esercice Echange Cellulaire PDFDocument5 pagesEsercice Echange Cellulaire PDFMael Njongang100% (1)
- p1 EleveDocument14 pagesp1 Eleveزكرياء بنحيرتPas encore d'évaluation
- Membranes UFDocument30 pagesMembranes UFOliver FrerePas encore d'évaluation
- Cour SVT TleDDocument122 pagesCour SVT TleDNdjidama youssoufa0% (1)
- Physiologie Des Voies Lacrymales DR LinaDocument34 pagesPhysiologie Des Voies Lacrymales DR Linaaymenkchaou31Pas encore d'évaluation
- Cours-GP-GPMAT-M1-Phénomènes de Surface Et Interfaces JijelDocument12 pagesCours-GP-GPMAT-M1-Phénomènes de Surface Et Interfaces JijelAymen BouguelaaPas encore d'évaluation
- LestechniquesmembranairesDocument44 pagesLestechniquesmembranairesAbir MabroukPas encore d'évaluation
- OsmométrieDocument13 pagesOsmométriemoha khb50% (4)
- Loriane Allard 300219504 A06 OsmoseDocument27 pagesLoriane Allard 300219504 A06 Osmoselorianeallard7Pas encore d'évaluation
- TP 03 Losmose Et La Pérmeabilite CellulaireDocument2 pagesTP 03 Losmose Et La Pérmeabilite Cellulairekim namjoonPas encore d'évaluation
- Étude - . .. - Des': Mesure Et Transpo Vts Solides I. 1:,, en Suspension Au Cameroun . .' YDocument43 pagesÉtude - . .. - Des': Mesure Et Transpo Vts Solides I. 1:,, en Suspension Au Cameroun . .' YdjouPas encore d'évaluation
- Cours Echanges Deau - Osmose FDocument5 pagesCours Echanges Deau - Osmose FDammak S.Pas encore d'évaluation
- TD Tle D Svteehb Échanges CellulairesDocument8 pagesTD Tle D Svteehb Échanges Cellulairesdrabodiane3Pas encore d'évaluation
- MDPPPPDocument9 pagesMDPPPPInes DaoudiPas encore d'évaluation
- ChromatographieDocument14 pagesChromatographieadilouche100% (1)
- Les Echanges Entre Les Plantes Et Le Sol CoursDocument16 pagesLes Echanges Entre Les Plantes Et Le Sol Courshamza manafPas encore d'évaluation
- Les Echanges Entre Les Plantes Et Le Sol CoursDocument16 pagesLes Echanges Entre Les Plantes Et Le Sol CoursAli AliPas encore d'évaluation
- Cours+4 PDF 22Document29 pagesCours+4 PDF 22Aymen ChougradPas encore d'évaluation
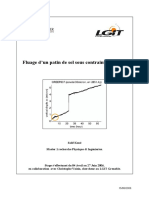
- Memoire Master 1 Physique & Ingenierie Kone Salif Lgit 2006 Universite Grenoble1Document22 pagesMemoire Master 1 Physique & Ingenierie Kone Salif Lgit 2006 Universite Grenoble1Kone SalifPas encore d'évaluation
- Application de La Pression Osmotique PDFDocument4 pagesApplication de La Pression Osmotique PDFZied BouabenePas encore d'évaluation
- La Stabilite Des Mousse LiquideDocument31 pagesLa Stabilite Des Mousse LiquideMessaoud Ait El HadiPas encore d'évaluation
- Cours Et Exercices Corrigés MDFDocument110 pagesCours Et Exercices Corrigés MDFchadou salem100% (4)
- TD 5 Phénomène de DiffusionDocument2 pagesTD 5 Phénomène de DiffusionBelinda Dancheu100% (1)
- Exposé 55Document14 pagesExposé 55clevickossoko28Pas encore d'évaluation
- Procédés D'adsorptionDocument6 pagesProcédés D'adsorptionkrim gasmiPas encore d'évaluation
- CCMDocument1 pageCCMnadpharm13Pas encore d'évaluation
- S-47751-4 BICHATE Anatomie Generale Tomo IV PDFDocument426 pagesS-47751-4 BICHATE Anatomie Generale Tomo IV PDFalsbemPas encore d'évaluation
- TP3 Céramique - Masse Volumique, Frittage Et MicrostructuresDocument15 pagesTP3 Céramique - Masse Volumique, Frittage Et MicrostructuresazertPas encore d'évaluation
- Essai Œdométrique Compressibilité, Perméabilité: Estp Laboratoire de GéotechniqueDocument7 pagesEssai Œdométrique Compressibilité, Perméabilité: Estp Laboratoire de GéotechniquebobPas encore d'évaluation
- Article de ColmatageDocument8 pagesArticle de ColmatageZakia ZakiaPas encore d'évaluation
- Détecteurs de particules: Les Grands Articles d'UniversalisD'EverandDétecteurs de particules: Les Grands Articles d'UniversalisPas encore d'évaluation
- Chromatographie: Les Grands Articles d'UniversalisD'EverandChromatographie: Les Grands Articles d'UniversalisPas encore d'évaluation
- Œil humain: Les Grands Articles d'UniversalisD'EverandŒil humain: Les Grands Articles d'UniversalisPas encore d'évaluation
- Membranes cellulaires: Les Grands Articles d'UniversalisD'EverandMembranes cellulaires: Les Grands Articles d'UniversalisPas encore d'évaluation
- Physiologie CardiovasculaireDocument76 pagesPhysiologie CardiovasculaireDavid FMPas encore d'évaluation
- ITEM 184p - ASTHMEDocument5 pagesITEM 184p - ASTHMESofia RhellabPas encore d'évaluation
- 03 1 PDFDocument4 pages03 1 PDFAbdelkrim AbdelkrimPas encore d'évaluation
- CAT Devant LeucorrhéeDocument27 pagesCAT Devant Leucorrhéeanis anis0% (1)
- Le Serum Sale Hypertonique en 2004Document8 pagesLe Serum Sale Hypertonique en 2004Alain SoucotPas encore d'évaluation
- ShigellosesDocument83 pagesShigellosesBoubacar BarryPas encore d'évaluation
- Pleurésie Non Purulente Bambey - Version LongueDocument59 pagesPleurésie Non Purulente Bambey - Version LongueChaimae TahiriPas encore d'évaluation
- EXERCICESDocument6 pagesEXERCICESثصلصلqsfcqfPas encore d'évaluation
- Maladies Rares en Medecine D'urgence by Yann-Erick Claessens, Luc Mouthon (Auth.)Document425 pagesMaladies Rares en Medecine D'urgence by Yann-Erick Claessens, Luc Mouthon (Auth.)Elena EllaPas encore d'évaluation
- Foie - Définition, Schéma, Anatomie Et RôleDocument5 pagesFoie - Définition, Schéma, Anatomie Et RôleNarath LongPas encore d'évaluation
- Syndrome de VOLKMAN Cours Key 2020Document4 pagesSyndrome de VOLKMAN Cours Key 2020Amï NãPas encore d'évaluation
- LipothymiesetsyncopesDocument6 pagesLipothymiesetsyncopesYoussouf OuedraogoPas encore d'évaluation
- Anatomie-Physiologie de L'appareil Respiratoire PDFDocument72 pagesAnatomie-Physiologie de L'appareil Respiratoire PDFYassine Drh100% (1)
- Le Bilan Biologique en OdontostomatologieDocument41 pagesLe Bilan Biologique en Odontostomatologieimene bougheziouPas encore d'évaluation
- A Lire - Cancer - L'éviter Ou Le Soigner Sans Mutilation. - La Table RondeDocument5 pagesA Lire - Cancer - L'éviter Ou Le Soigner Sans Mutilation. - La Table RondeCatherine DelamaisonPas encore d'évaluation
- Les Toxines À Tropisme MembranaireDocument9 pagesLes Toxines À Tropisme Membranairesoumi soumiaPas encore d'évaluation
- Lithiase Urinaire PDFDocument22 pagesLithiase Urinaire PDFsyrille clerienPas encore d'évaluation
- Système MusculaireDocument7 pagesSystème MusculaireAmine WlPas encore d'évaluation
- Llopis Benoît. Thèse D'exercice de Pharmacie (UPJV)Document120 pagesLlopis Benoît. Thèse D'exercice de Pharmacie (UPJV)IbrahimPas encore d'évaluation
- Fichier Produit 3105Document9 pagesFichier Produit 3105Darryl LouisPas encore d'évaluation
- IdrissDocument37 pagesIdrissDominique Ilunga mbambiPas encore d'évaluation
- Etude Épidémiologique de La Drépanocytose Dans La Région de ConstantineDocument55 pagesEtude Épidémiologique de La Drépanocytose Dans La Région de ConstantineSimon KubelezagaPas encore d'évaluation
- Dépistage de Tuberculose ç-1Document18 pagesDépistage de Tuberculose ç-1Samira Moutaoikkil BasskarPas encore d'évaluation
- Notice Debridat 100mg Comp - Pelli. B 20Document4 pagesNotice Debridat 100mg Comp - Pelli. B 20son.600Pas encore d'évaluation
- MedecinetropicaleDocument16 pagesMedecinetropicalecammganeshPas encore d'évaluation
- Le - Tissus Osseaux Et Les OsDocument89 pagesLe - Tissus Osseaux Et Les OsNettah MarouaPas encore d'évaluation
- Wepik Approche Clinique Et Gestion de La Mastoidite Aigue Une Etude Approfondie 20231203222155rUSXDocument18 pagesWepik Approche Clinique Et Gestion de La Mastoidite Aigue Une Etude Approfondie 20231203222155rUSXAsmaa MorsliPas encore d'évaluation
- Introduction À La MicrobiologieDocument42 pagesIntroduction À La MicrobiologieRaïs MahoungouPas encore d'évaluation
- Syllabus CARDIO BA3 2021-2022Document117 pagesSyllabus CARDIO BA3 2021-2022Alexandre BerchouchiPas encore d'évaluation
- Achondroplasie FR FR HAN ORPHA15Document5 pagesAchondroplasie FR FR HAN ORPHA15youssef EL KHAYATIPas encore d'évaluation