Vous aimerez peut-être aussi
- Immunologie PDFDocument95 pagesImmunologie PDFdjassa djassaPas encore d'évaluation
- Sclérose en Plaques - NetterDocument9 pagesSclérose en Plaques - NetterS. DonyaPas encore d'évaluation
- 2016 Les Accidents Transfusionnels PDFDocument42 pages2016 Les Accidents Transfusionnels PDFMohamed Ali MtibaaPas encore d'évaluation
- Cellules Et Organes ImmunitairesDocument28 pagesCellules Et Organes ImmunitairesInes Ben HniaPas encore d'évaluation
- 5 - Physiologie de La SomésthésieDocument11 pages5 - Physiologie de La SomésthésieEmelie MelodyPas encore d'évaluation
- QCM Topographie PelvienneDocument6 pagesQCM Topographie PelvienneYoBjj100% (2)
- Résidents HEMATOPOIESEDocument38 pagesRésidents HEMATOPOIESEMeshoub Soumia100% (1)
- Les Cellules Lymphoides 08Document36 pagesLes Cellules Lymphoides 08YASMEROPas encore d'évaluation
- Anatomie 08Document3 pagesAnatomie 08Hamza Adri100% (1)
- 03 Physiologie Du GR 2020 PDFDocument22 pages03 Physiologie Du GR 2020 PDFNabyl BektachePas encore d'évaluation
- Les Tissus Conjonctifs Et Musculaires PCEM1 - 2021Document56 pagesLes Tissus Conjonctifs Et Musculaires PCEM1 - 2021Wala DallaliPas encore d'évaluation
- Anatomie SchémaDocument57 pagesAnatomie Schémakarim benhaddouPas encore d'évaluation
- L5-Processus Celllulaires de Défense Contre Les Agents InfectieuxDocument9 pagesL5-Processus Celllulaires de Défense Contre Les Agents InfectieuxRãsí ÃckėrmānPas encore d'évaluation
- Lymphocytes T Et BDocument48 pagesLymphocytes T Et BTul IpPas encore d'évaluation
- Le Systeme Du ComplementDocument8 pagesLe Systeme Du ComplementAymenPas encore d'évaluation
- Antibiotherapie NewDocument62 pagesAntibiotherapie NewGÉNÉRAL TECHPas encore d'évaluation
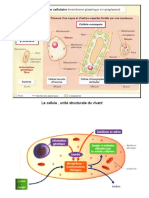
- Cyto1an-Systeme EndomembranaireDocument82 pagesCyto1an-Systeme EndomembranaireAdam FellousPas encore d'évaluation
- Le CMHDocument9 pagesLe CMHHabib DarouichePas encore d'évaluation
- Anomalies Chromosomiques DR Maayoufi PDFDocument9 pagesAnomalies Chromosomiques DR Maayoufi PDFhalaPas encore d'évaluation
- Le Système Du ComplémentDocument9 pagesLe Système Du Complémenttara mourad100% (1)
- Hyperlymphocytose Version FraDocument36 pagesHyperlymphocytose Version Framohmoh tiki takaPas encore d'évaluation
- Association HLA2021Document37 pagesAssociation HLA2021Mohamed AjroudPas encore d'évaluation
- Immunoglobulines CoursDocument11 pagesImmunoglobulines CoursAloui RayenPas encore d'évaluation
- Anapath3an-Processus Inflammatoire2022benahseneDocument10 pagesAnapath3an-Processus Inflammatoire2022benahseneouss said100% (1)
- Chapitre II SpermatogenèseDocument6 pagesChapitre II SpermatogenèseDel HadPas encore d'évaluation
- Lymphopoïèse Lymphocytes DR BOUHSANEDocument6 pagesLymphopoïèse Lymphocytes DR BOUHSANENadera KessarPas encore d'évaluation
- Cours 9 Caryotype 2020Document5 pagesCours 9 Caryotype 2020Farah B. BtoushPas encore d'évaluation
- ImmunitéDocument31 pagesImmunitélahlou Lh100% (1)
- Eucaryotes Et Virologie (Version 2)Document24 pagesEucaryotes Et Virologie (Version 2)coursenligne.lille1100% (3)
- Chap 2Document5 pagesChap 2Ghribi SanaPas encore d'évaluation
- Introduction Générale À LimmunologieDocument67 pagesIntroduction Générale À LimmunologieNabil holmes100% (1)
- Civd Uz 2022Document7 pagesCivd Uz 2022Be ExpensivePas encore d'évaluation
- Sérologie de L'auto-ImmunitéDocument18 pagesSérologie de L'auto-Immunitéasmahane mohammediPas encore d'évaluation
- Organogénèse Diapos - Cours Maïeutique JOGUET P1 - UE8 - UE11 de L'université Des Antilles Et de La GuadeloupeDocument20 pagesOrganogénèse Diapos - Cours Maïeutique JOGUET P1 - UE8 - UE11 de L'université Des Antilles Et de La GuadeloupeUniversité des AntillesPas encore d'évaluation
- 1 Physiologie Du NeuroneDocument26 pages1 Physiologie Du NeuroneSahouin Claude MedardPas encore d'évaluation
- Exploration de L'hémostaseDocument18 pagesExploration de L'hémostaseKhaoula SahtouriPas encore d'évaluation
- Medecine-Intro Genetique MonohybridismeDocument23 pagesMedecine-Intro Genetique MonohybridismedzsergioPas encore d'évaluation
- HémoglobineDocument32 pagesHémoglobineaminePas encore d'évaluation
- Cours de LhématopoïèseDocument15 pagesCours de LhématopoïèseIkbel WederniPas encore d'évaluation
- Histologie SB Med Phar CM 2020Document66 pagesHistologie SB Med Phar CM 2020yvesPas encore d'évaluation
- 1-Troubles Du Métabolisme Des Glucides PDFDocument5 pages1-Troubles Du Métabolisme Des Glucides PDFAisSa KeRrøùùm100% (1)
- 3 SomesthésieDocument25 pages3 SomesthésieSahouin Claude MedardPas encore d'évaluation
- Immuno 03. LyT Khadidja FerdjDocument8 pagesImmuno 03. LyT Khadidja Ferdjmaya mhdPas encore d'évaluation
- Syst CardiocirculatoireDocument187 pagesSyst CardiocirculatoireAsma KalfallahPas encore d'évaluation
- Caryotype DR HammoudaDocument5 pagesCaryotype DR HammoudahalaPas encore d'évaluation
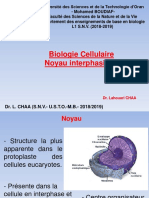
- Cours Biologie Cellulaire Noyau Interphasique 2018 2019Document11 pagesCours Biologie Cellulaire Noyau Interphasique 2018 2019amel bekkayePas encore d'évaluation
- Cours - 2 Hématopoïèse PDFDocument15 pagesCours - 2 Hématopoïèse PDFMohamed Amine BakariPas encore d'évaluation
- Page82 11. Polynucleaires Monocytes Et MacrophagesDocument13 pagesPage82 11. Polynucleaires Monocytes Et Macrophageskaka lotyPas encore d'évaluation
- Les Fièvres 2023Document59 pagesLes Fièvres 2023CoulibalyPas encore d'évaluation
- Physio SOMESTHESIEDocument25 pagesPhysio SOMESTHESIEEsperancePas encore d'évaluation
- GRANULOPOIESEDocument48 pagesGRANULOPOIESEYingra BaraPas encore d'évaluation
- Chaine RespiratoireDocument1 pageChaine RespiratoireayoubborombaPas encore d'évaluation
- 7-Pa Et Sa RegulationDocument4 pages7-Pa Et Sa RegulationImene BenzianePas encore d'évaluation
- A. Embryo 06 Deuxième Semaine Du Développement EmbryonnaireDocument12 pagesA. Embryo 06 Deuxième Semaine Du Développement EmbryonnaireILHEM RAMDAPas encore d'évaluation
- Tout La Physio en QCM ???Document226 pagesTout La Physio en QCM ???ikijissamPas encore d'évaluation
- Granulopoïèse Et Sa RégulationDocument24 pagesGranulopoïèse Et Sa RégulationMor Ngom100% (1)
- Canaux CalciqueDocument4 pagesCanaux CalciqueAchraf HakkPas encore d'évaluation
- Tissu OsseuxDocument39 pagesTissu OsseuxBMA-medecine100% (2)
- Cours IV Tolérance Immunitaire Et Auto ImmunitéDocument13 pagesCours IV Tolérance Immunitaire Et Auto ImmunitétinaPas encore d'évaluation
- Hemostase Primaire 2018Document26 pagesHemostase Primaire 2018fifi fifiPas encore d'évaluation
- 02 cmh2021zemouliDocument14 pages02 cmh2021zemoulidanger inflammablePas encore d'évaluation
- Immunologie. CMHDocument8 pagesImmunologie. CMHËl kíng čhrístíän MükänïrwäPas encore d'évaluation
- Les ImmunoglobulinesDocument9 pagesLes Immunoglobulinestara mouradPas encore d'évaluation
- Les AntigènesDocument4 pagesLes Antigènestara mouradPas encore d'évaluation
- Les Cytokines 2019 2020Document7 pagesLes Cytokines 2019 2020tara mouradPas encore d'évaluation
- La Réaction InflammatoireDocument8 pagesLa Réaction Inflammatoiretara mouradPas encore d'évaluation
- Sein Echographiemammaire SorDocument6 pagesSein Echographiemammaire Sortara mouradPas encore d'évaluation
- Seno Echo SeinDocument2 pagesSeno Echo Seintara mouradPas encore d'évaluation
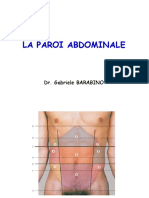
- Paroi Abdominale1Document17 pagesParoi Abdominale1Fatima Zahra Rahim ArchiPas encore d'évaluation
- Correction Zoologie Examen BlancDocument2 pagesCorrection Zoologie Examen Blancyoussouf traore100% (2)
- Devoir01 t10Document4 pagesDevoir01 t10Compaore Abdoul DesrachPas encore d'évaluation
- 1 La Pratique de L'allaitement Maternel - D - SoDocument93 pages1 La Pratique de L'allaitement Maternel - D - SoAMINAPas encore d'évaluation
- BA Prof CoursDocument224 pagesBA Prof Coursbenjelloun ikramPas encore d'évaluation
- Immunité de La Cavité BuccaleDocument12 pagesImmunité de La Cavité Buccaleمحاضرات علمية0% (1)
- 2-Le Metabolisme Des AaDocument3 pages2-Le Metabolisme Des AaoutsiderPas encore d'évaluation
- Carte Mental SNDocument1 pageCarte Mental SNsimon.praudayraudPas encore d'évaluation
- Physiologie de L'appareil Respiratoire - PR FaroudyDocument14 pagesPhysiologie de L'appareil Respiratoire - PR FaroudyMarouane bouhabsPas encore d'évaluation
- 04 - 45 - 02léiomyome Variantes Et RemaniementsDocument46 pages04 - 45 - 02léiomyome Variantes Et RemaniementsjelenaPas encore d'évaluation
- Interactions Ligand - Recepteur 2023-2024Document29 pagesInteractions Ligand - Recepteur 2023-2024khaldiniheddoc23Pas encore d'évaluation
- Zabr JT5 ActesDocument110 pagesZabr JT5 Acteskarim sbihiPas encore d'évaluation
- Dragonfury Tome 4Document246 pagesDragonfury Tome 4Charlotte CornuPas encore d'évaluation
- 19.troubles Hydro ÉlectrolytiqueDocument16 pages19.troubles Hydro ÉlectrolytiqueMohamed Boussif100% (1)
- ORTH 2019 GiraudDocument78 pagesORTH 2019 GiraudlylyPas encore d'évaluation
- Innervation FaceDocument8 pagesInnervation FaceFree CoursesPas encore d'évaluation
- Hemostase Primaire 2018Document26 pagesHemostase Primaire 2018fifi fifiPas encore d'évaluation
- TB Hydratation PDFDocument40 pagesTB Hydratation PDFAdognon Jaquet DeJésusPas encore d'évaluation
- Bio Ani VertebresDocument86 pagesBio Ani VertebresmaomaochongPas encore d'évaluation
- Chapitre 02 Les Tissus SecondairesDocument5 pagesChapitre 02 Les Tissus SecondairesThe VoidPas encore d'évaluation
- Crce & Dévpt - L3 - UMAN - ET - 2019Document159 pagesCrce & Dévpt - L3 - UMAN - ET - 2019Koffi WilfriedPas encore d'évaluation
- SNtestDocument3 pagesSNtestMagouePas encore d'évaluation
- Stock LesPdf Examens BAC Comores Sujet 2011 Comores Sujet D Science (SVT) Bac 2011Document4 pagesStock LesPdf Examens BAC Comores Sujet 2011 Comores Sujet D Science (SVT) Bac 2011Ahmed100% (1)
- Chap 4 EnzymesDocument38 pagesChap 4 Enzymesflo bilPas encore d'évaluation
- TD n2: La Soif Et La Faim: DR Ghalem MeriemDocument41 pagesTD n2: La Soif Et La Faim: DR Ghalem Meriemfatimazahraaelhammioui0Pas encore d'évaluation
- 2015 11 Protocole Parcours de Soins VFDocument12 pages2015 11 Protocole Parcours de Soins VFzouiche KhadidjaPas encore d'évaluation
- BiophysiqueDocument28 pagesBiophysiqueYannick DsprbsPas encore d'évaluation
- Devoir N1 - Semestre 1 - SVT 1AC Modele Word 1Document2 pagesDevoir N1 - Semestre 1 - SVT 1AC Modele Word 1Soufiane KarchaliPas encore d'évaluation
- Systeme Digestif ResumeDocument2 pagesSysteme Digestif ResumehelloPas encore d'évaluation