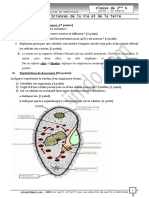
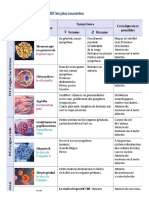
Vous aimerez peut-être aussi
- Annales SVT 3eDocument74 pagesAnnales SVT 3eJosoa TOKINJANAHARYPas encore d'évaluation
- La Diète 3 Semaines PDF GratuitDocument49 pagesLa Diète 3 Semaines PDF GratuitBrian Flatt50% (2)
- Annales SVT Tle DDocument127 pagesAnnales SVT Tle DJunior NoulaPas encore d'évaluation
- Annales SVT Tle d508Document245 pagesAnnales SVT Tle d508SIRIKI YEOPas encore d'évaluation
- Exercice de GénétiqueDocument69 pagesExercice de GénétiqueOumarou KontaPas encore d'évaluation
- La Division CellulaireDocument30 pagesLa Division Cellulaireselma torresPas encore d'évaluation
- Théorie des temps grammaticaux fondée sur les traits pertinents temporelsD'EverandThéorie des temps grammaticaux fondée sur les traits pertinents temporelsPas encore d'évaluation
- SVT Sébikotane 1ère DevoirDocument2 pagesSVT Sébikotane 1ère DevoirAminata89% (9)
- Le langage au bout des doigts: Les liens fonctionnels entre la motricité et le langageD'EverandLe langage au bout des doigts: Les liens fonctionnels entre la motricité et le langagePas encore d'évaluation
- Des chemins de la réussite: La gestion mentale en renfort des classes de sciencesD'EverandDes chemins de la réussite: La gestion mentale en renfort des classes de sciencesPas encore d'évaluation
- OsmoseDocument17 pagesOsmosetomeyumPas encore d'évaluation
- Genetique Medicale GUGDocument155 pagesGenetique Medicale GUGanaPas encore d'évaluation
- Séquençage d'ADNDocument27 pagesSéquençage d'ADNFati100% (11)
- 2ème - physio.LA TRANSMISSION SYNAPTIQUE PDFDocument37 pages2ème - physio.LA TRANSMISSION SYNAPTIQUE PDFكوردورلي الحاجPas encore d'évaluation
- EPREUVE DE SVTEEHB - Docx PD Sequence IDocument5 pagesEPREUVE DE SVTEEHB - Docx PD Sequence IElie ZRAPas encore d'évaluation
- TD2BMS6 PDFDocument88 pagesTD2BMS6 PDFAmine BeninePas encore d'évaluation
- Supports SVT PDFDocument38 pagesSupports SVT PDFzineb EZZINEPas encore d'évaluation
- SVTEEHB 1ère D Probatoire Blanc 2 Mai 2023Document4 pagesSVTEEHB 1ère D Probatoire Blanc 2 Mai 2023Loic UlrichPas encore d'évaluation
- Tigp's Cc3 Svteehb TD Vac2021Document4 pagesTigp's Cc3 Svteehb TD Vac2021Antoinette AdjakpleyPas encore d'évaluation
- Bio 110Document148 pagesBio 110stones bankPas encore d'évaluation
- 1ere D Probat. Blanc 2004-2005Document9 pages1ere D Probat. Blanc 2004-2005tohamyebgafernandjuniorulrichPas encore d'évaluation
- Spé SVTDocument8 pagesSpé SVTanissaPas encore d'évaluation
- Renf MitoseDocument4 pagesRenf MitoseIsmaila TraoréPas encore d'évaluation
- 2022 Syllabus Bio 110 Octobre 22 MoodleDocument4 pages2022 Syllabus Bio 110 Octobre 22 MoodleAmèvi Mawugno AGBOVIPas encore d'évaluation
- TIGp's - TPE 1 - SVTEEHB - T DDocument25 pagesTIGp's - TPE 1 - SVTEEHB - T DLandry Tenga100% (1)
- SVTEEHB - LYCEE DE OULARGO - SÉQUENCE N°1 CLASSE TLE D CAMEROUNDocument3 pagesSVTEEHB - LYCEE DE OULARGO - SÉQUENCE N°1 CLASSE TLE D CAMEROUNlatifamoyopo9Pas encore d'évaluation
- BIO5064 GuideDocument154 pagesBIO5064 GuideYacine YacinePas encore d'évaluation
- Cours - Cytologie Histologie - Module1 Séquuences1 6 PDFDocument117 pagesCours - Cytologie Histologie - Module1 Séquuences1 6 PDFHenry Zeemann100% (1)
- WMD1120P: Biologie Générale Et Approche Expérimentale de La Biologie (Partim Biologie Générale)Document2 pagesWMD1120P: Biologie Générale Et Approche Expérimentale de La Biologie (Partim Biologie Générale)Honoré IrengePas encore d'évaluation
- 2022 09 Sep20 Introduction Du CoursDocument17 pages2022 09 Sep20 Introduction Du CoursNouhoum TraoréPas encore d'évaluation
- 1.BCB-05 Membrane PlasmiqueDocument39 pages1.BCB-05 Membrane PlasmiqueMastan AttouchePas encore d'évaluation
- 1-Le Nucléotide Cytidine Monophosphate Est Constitué:: D. +cytosine + Acide PhosphoriqueDocument4 pages1-Le Nucléotide Cytidine Monophosphate Est Constitué:: D. +cytosine + Acide PhosphoriqueSimonPas encore d'évaluation
- Le Facilitateur PD Correction 1Document61 pagesLe Facilitateur PD Correction 1Louis Yannick EssombaPas encore d'évaluation
- Questions de Lexamen 1Document39 pagesQuestions de Lexamen 1dhouha14ghorbelPas encore d'évaluation
- Bac Science NatuDocument79 pagesBac Science NatuLine Hustling DollarPas encore d'évaluation
- DS1 22Document3 pagesDS1 22rahma larianiPas encore d'évaluation
- Devoir de Synthèse N°3 Avec Correction 2010 2011 (Said Mounir)Document8 pagesDevoir de Synthèse N°3 Avec Correction 2010 2011 (Said Mounir)Sana Ghribi100% (1)
- Dre-S CRPS SVT 1D 2023-2024Document3 pagesDre-S CRPS SVT 1D 2023-2024gilberthouzoukePas encore d'évaluation
- Cours Bio Cell 2011-EtudiantsDocument23 pagesCours Bio Cell 2011-EtudiantsSalma BabourPas encore d'évaluation
- Cours Bio Cell 2011-EtudiantsDocument23 pagesCours Bio Cell 2011-EtudiantsAmï NãPas encore d'évaluation
- Cours Bio Cell 2011-EtudiantsDocument23 pagesCours Bio Cell 2011-EtudiantsAmï NãPas encore d'évaluation
- Cours Bio Cell 2011-EtudiantsDocument23 pagesCours Bio Cell 2011-EtudiantsRami NadjmPas encore d'évaluation
- Cours Bio Cell 2011-EtudiantsDocument23 pagesCours Bio Cell 2011-EtudiantsARMAND KUIPOUPas encore d'évaluation
- TD N 1 Biologie MoléculaireDocument6 pagesTD N 1 Biologie Moléculairefirdaoussaoud22Pas encore d'évaluation
- Introduction A La Biologie Cellulaire Et Moleculaire ImprimeDocument215 pagesIntroduction A La Biologie Cellulaire Et Moleculaire ImprimeMc UltraHopePas encore d'évaluation
- Svteehb Pd1 TD 2019-2020Document3 pagesSvteehb Pd1 TD 2019-2020Oreol OrealPas encore d'évaluation
- 16-19 BioDocument144 pages16-19 BiowongxiuhoPas encore d'évaluation
- SVTEEHB TleA Seq2 College Eteudem 2020 2021Document2 pagesSVTEEHB TleA Seq2 College Eteudem 2020 2021Gradur ScarfacePas encore d'évaluation
- REVISION BIOS 202: Numéro 2 Exercice 1: Dans Un Récipient Contenant 1 Litre D'eau, On AjouteDocument2 pagesREVISION BIOS 202: Numéro 2 Exercice 1: Dans Un Récipient Contenant 1 Litre D'eau, On Ajouteyaa100% (1)
- TA 4 VraiDocument3 pagesTA 4 VraiSteeve LyonPas encore d'évaluation
- c s Les Archimedes d.s 2e Semestre Svt 1ière dDocument3 pagesc s Les Archimedes d.s 2e Semestre Svt 1ière dkpadedarius822Pas encore d'évaluation
- Biologie ST2S CR AtliersDocument10 pagesBiologie ST2S CR AtliersZa HraPas encore d'évaluation
- SERIE 1 cytologie 4AS 2023 _230512_214257Document2 pagesSERIE 1 cytologie 4AS 2023 _230512_214257abdoumicrobio75Pas encore d'évaluation
- 2 NDDocument2 pages2 NDAubin TcheunouPas encore d'évaluation
- FA La CelluleDocument7 pagesFA La Cellulenourabakayokoo10Pas encore d'évaluation
- Epreuve de Svteehb PD 1er SeqDocument3 pagesEpreuve de Svteehb PD 1er SeqElie ZRAPas encore d'évaluation
- Syllabus Biologie Cellulaire- PHARMACIE_License 1_2022-2023Document2 pagesSyllabus Biologie Cellulaire- PHARMACIE_License 1_2022-2023yehiyadicko1Pas encore d'évaluation
- Devoir Corrige de Synthese N3 SVT 2eme SDocument8 pagesDevoir Corrige de Synthese N3 SVT 2eme Schafitalbi72Pas encore d'évaluation
- Devoir Corrigé de Synthèse N°3 - SVT - 2ème Sciences (2010-2011) MR Said MounirDocument8 pagesDevoir Corrigé de Synthèse N°3 - SVT - 2ème Sciences (2010-2011) MR Said Mounirelyes mleykiPas encore d'évaluation
- Thème I.S2.Cours - ÉlèvesdocxDocument4 pagesThème I.S2.Cours - ÉlèvesdocxLivre BDPas encore d'évaluation
- Capture D'écran . 2022-11-09 À 23.26.41Document6 pagesCapture D'écran . 2022-11-09 À 23.26.41Bouchra KriPas encore d'évaluation
- TD Tle D Svteehb Échanges CellulairesDocument8 pagesTD Tle D Svteehb Échanges Cellulairesdrabodiane3Pas encore d'évaluation
- LSV1 CM I 21Document36 pagesLSV1 CM I 21kirito 83Pas encore d'évaluation
- Biologie-Cellulaire - LIVRE de 30 FicheDocument165 pagesBiologie-Cellulaire - LIVRE de 30 FicheRahma LionnePas encore d'évaluation
- Biologie 12 EmeDocument48 pagesBiologie 12 EmeSawoulou celest DopavoguiPas encore d'évaluation
- CSVT-102-La Replication Cycle CellulaireDocument5 pagesCSVT-102-La Replication Cycle CellulaireJean-Pierre PolnareffPas encore d'évaluation
- 1SVT 2ndeDocument29 pages1SVT 2ndeJulie MPas encore d'évaluation
- Annale - BiologieDocument71 pagesAnnale - BiologieDesakoBEATSPas encore d'évaluation
- La Demarche PedagogiqueDocument2 pagesLa Demarche Pedagogiquezineb EZZINEPas encore d'évaluation
- Sciences Naturelles: AnnalesDocument182 pagesSciences Naturelles: Annaleszineb EZZINEPas encore d'évaluation
- C - BIOLOGIE@TleD - II - LES MOUVEMENTS ET LES ECHANGES CELLULAIRES PDFDocument23 pagesC - BIOLOGIE@TleD - II - LES MOUVEMENTS ET LES ECHANGES CELLULAIRES PDFzineb EZZINEPas encore d'évaluation
- Les Caracteristiques Des Ist Les Plus Courantes PDFDocument2 pagesLes Caracteristiques Des Ist Les Plus Courantes PDFzineb EZZINEPas encore d'évaluation
- Atelier2 Scenarisation Pédagogique SVTDocument23 pagesAtelier2 Scenarisation Pédagogique SVTzineb EZZINEPas encore d'évaluation
- 2 Serie td2 Les Echanges Cellulaires 2016 2017Document2 pages2 Serie td2 Les Echanges Cellulaires 2016 2017Stephan Effim67% (3)
- Guide Prof-Al Moufid SVT-2AC - 22 Juillet 2019Document83 pagesGuide Prof-Al Moufid SVT-2AC - 22 Juillet 2019zineb EZZINEPas encore d'évaluation
- Choc Séptique 2020 Réeele PDFDocument20 pagesChoc Séptique 2020 Réeele PDFHassan BerdaiPas encore d'évaluation
- C2 Les AntifongiquesDocument14 pagesC2 Les AntifongiquesAlexandre Ouffoue100% (2)
- TD 3 Le CytosqueletteDocument7 pagesTD 3 Le CytosqueletteĐî ŅæPas encore d'évaluation
- PCR en Temps Réel en Virologie: Enquête Et RésuméDocument14 pagesPCR en Temps Réel en Virologie: Enquête Et RésuméStarix UkrPas encore d'évaluation
- SUPPORT UE1B-KREJBICH-Relation Structure Fonction Des Biomolécules-19.09.22Document60 pagesSUPPORT UE1B-KREJBICH-Relation Structure Fonction Des Biomolécules-19.09.22Yann legendrePas encore d'évaluation
- Cours - DJERMOUNE Lynda - Biochimie AlimentaireDocument107 pagesCours - DJERMOUNE Lynda - Biochimie AlimentairessalahPas encore d'évaluation
- Liberation de L Energie Emmagasinee Dans La Matiere Organique CoursDocument12 pagesLiberation de L Energie Emmagasinee Dans La Matiere Organique CourspdfPas encore d'évaluation
- Fiche Calcul de La Charge Dun PeptideDocument4 pagesFiche Calcul de La Charge Dun PeptidePhy MedPas encore d'évaluation
- 2.2 BrdU CorrecDocument2 pages2.2 BrdU CorrecReyzPas encore d'évaluation
- Devoir de Première S N°2 - Type II2Document3 pagesDevoir de Première S N°2 - Type II2Nihed BelhadjiPas encore d'évaluation
- Bioinformatique 3ème Licence Microbiologie 27 05 2018Document15 pagesBioinformatique 3ème Licence Microbiologie 27 05 2018BENYAHIA NESRINEPas encore d'évaluation
- Biochimie - Introduction Aux TechniquesDocument5 pagesBiochimie - Introduction Aux TechniquesmolandPas encore d'évaluation
- Cour 4Document20 pagesCour 4sino spagoPas encore d'évaluation
- SVT DM 27-03Document2 pagesSVT DM 27-03Camille BerthetPas encore d'évaluation
- Transgenèse Végétale Et ApplicationsDocument16 pagesTransgenèse Végétale Et Applicationscoursenligne.lille1100% (13)
- Toxines Et Transferts IoniquesDocument180 pagesToxines Et Transferts IoniquesfaiahPas encore d'évaluation
- Bioch S2Document36 pagesBioch S2Eduard SimaderPas encore d'évaluation
- Acidose LactiqueDocument47 pagesAcidose Lactiqueيارب اسالك الجنةPas encore d'évaluation
- 1.2.2-Exercices OsidesDocument2 pages1.2.2-Exercices OsidesLawrence Mundene-timothee100% (2)
- 101-901-RE Dev 01GDocument11 pages101-901-RE Dev 01GgabnellyPas encore d'évaluation
- Sciences (SVT) Synthèse 2 Ème Seondaire Science 6580 Other44 EduluxDocument4 pagesSciences (SVT) Synthèse 2 Ème Seondaire Science 6580 Other44 EduluxKhawla SahliPas encore d'évaluation
- Evaluation Formative Partie 2 - CorrectionDocument12 pagesEvaluation Formative Partie 2 - Correctionineslina1709Pas encore d'évaluation
- Biochimie MicrobienneDocument29 pagesBiochimie Microbiennesino spagoPas encore d'évaluation