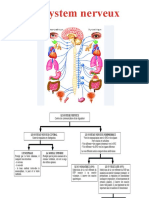
Vous aimerez peut-être aussi
- Neurobiologie: Les Grands Articles d'UniversalisD'EverandNeurobiologie: Les Grands Articles d'UniversalisPas encore d'évaluation
- Chapitre I Organisation Anatomofonctionnelle Du Systeme Nerveux0001Document19 pagesChapitre I Organisation Anatomofonctionnelle Du Systeme Nerveux0001Denisco NkemgnePas encore d'évaluation
- Chapitre I, II Et III de Physiologie Animale-2Document111 pagesChapitre I, II Et III de Physiologie Animale-2Talla NdaoPas encore d'évaluation
- Cours Neurones Synapses PDFDocument82 pagesCours Neurones Synapses PDFSofiane Salem100% (1)
- Physio 3Document22 pagesPhysio 3Sadi KoukouPas encore d'évaluation
- Cours de Neuroanatomie 2020-2021Document119 pagesCours de Neuroanatomie 2020-2021Dorcas BalalukaPas encore d'évaluation
- Physiopath Defmot MD 111-130Document18 pagesPhysiopath Defmot MD 111-130Sadi KoukouPas encore d'évaluation
- Chap II Tissu NerveuxDocument43 pagesChap II Tissu NerveuxbadamassimarwanePas encore d'évaluation
- Anatomie Et Physiologie Du Système Nerveux-1Document51 pagesAnatomie Et Physiologie Du Système Nerveux-1aurelien choupoPas encore d'évaluation
- Psychophysio PDF LMD 2015Document46 pagesPsychophysio PDF LMD 2015AliBenMessaoudPas encore d'évaluation
- TexteDocument3 pagesTexteDiallo OumarPas encore d'évaluation
- Syllabus Neurophysiologie Corrige-1Document131 pagesSyllabus Neurophysiologie Corrige-1Diawara Oumar100% (1)
- Neuroanat-2 0Document83 pagesNeuroanat-2 0seloua kaid100% (1)
- Exposer CerveauxDocument8 pagesExposer CerveauxKonan Armel KobenanPas encore d'évaluation
- Chap4 PapierDocument24 pagesChap4 PapierChristian abahPas encore d'évaluation
- Partie 4Document31 pagesPartie 4ndeye yarame diopPas encore d'évaluation
- 01 - Le Tissu NerveuxDocument13 pages01 - Le Tissu NerveuxNesrinePas encore d'évaluation
- CM4 - Vascularisation Et Voies Sensitivo-MotricesDocument14 pagesCM4 - Vascularisation Et Voies Sensitivo-Motricescharbelantoun2443Pas encore d'évaluation
- 01-Physilogie Du Tissu NerveuxDocument49 pages01-Physilogie Du Tissu NerveuxBook HunterPas encore d'évaluation
- Neurophysiologie 2Document49 pagesNeurophysiologie 2123kiwiPas encore d'évaluation
- Le System NerveuxDocument23 pagesLe System Nerveuxyamina kadiPas encore d'évaluation
- 1 Er Cour PhysiologieDocument13 pages1 Er Cour Physiologiederdiri imènePas encore d'évaluation
- NeurophysiologieDocument13 pagesNeurophysiologieCath Lemeunier100% (1)
- Neurophysiologie PDFDocument140 pagesNeurophysiologie PDFPascal Kasero0% (1)
- 1-Embryologie Du Systeme Nerveux - La Moelle ÉpinièreDocument17 pages1-Embryologie Du Systeme Nerveux - La Moelle ÉpinièreSist IAPas encore d'évaluation
- Ms Ebm Benhabib+BradaiDocument63 pagesMs Ebm Benhabib+BradaiDady LalaPas encore d'évaluation
- Le Tissu NerveuxDocument10 pagesLe Tissu NerveuxANDRIANOME tafitaPas encore d'évaluation
- Système Nerveux 1Document21 pagesSystème Nerveux 1Ranime Mekersi100% (1)
- Systeme Nerveux°0Document26 pagesSysteme Nerveux°0salifnchare16Pas encore d'évaluation
- Physiopathologie NeuroscienceDocument46 pagesPhysiopathologie NeuroscienceYas MiiinaaPas encore d'évaluation
- La Physiologie Du Systeme Nerveux Centrale DR GuedjatiDocument33 pagesLa Physiologie Du Systeme Nerveux Centrale DR GuedjatiMedMan6100% (2)
- Organisation AnatomofonctionnelleDocument95 pagesOrganisation AnatomofonctionnelleDenisco NkemgnePas encore d'évaluation
- Neurones Et Transmission SynaptiqueDocument6 pagesNeurones Et Transmission Synaptiquelouanne kloepferPas encore d'évaluation
- Système NerveuxDocument59 pagesSystème NerveuxBilal AliounePas encore d'évaluation
- NeurophysiologieDocument15 pagesNeurophysiologieaitbaali.oriflamePas encore d'évaluation
- Chapitre 3A Cours Sur Cerveau Et Mouvement VolontaireDocument3 pagesChapitre 3A Cours Sur Cerveau Et Mouvement Volontairealice.maranberPas encore d'évaluation
- Neuro Physio LogieDocument32 pagesNeuro Physio LogieAssia BsrPas encore d'évaluation
- Physiologie Du Nerf 2022 Mede UM6Document163 pagesPhysiologie Du Nerf 2022 Mede UM6Girl Ytb100% (1)
- Anatomie SN 2024Document53 pagesAnatomie SN 2024lwakwanasaidesPas encore d'évaluation
- MI3 Neurosciences Histologie Du Systeme Nerveux LavabreDocument17 pagesMI3 Neurosciences Histologie Du Systeme Nerveux LavabreDjallal HassaniPas encore d'évaluation
- 4eme Type Tissu NerveuxDocument36 pages4eme Type Tissu Nerveuxgroupe_svi3Pas encore d'évaluation
- 08 - Le Tissu NerveuxDocument7 pages08 - Le Tissu NerveuxlahlahazouPas encore d'évaluation
- Systeme NerveuxDocument82 pagesSysteme Nerveuxmohamedmahfoud198Pas encore d'évaluation
- 1a d1 Ue8 Cours 1 Developpement Du Systeme Nerveux Central-RoneoDocument14 pages1a d1 Ue8 Cours 1 Developpement Du Systeme Nerveux Central-RoneosyurmiPas encore d'évaluation
- Cahier TP1 Biomed 2022Document16 pagesCahier TP1 Biomed 2022Tr LwPas encore d'évaluation
- Le Tissu NerveuxDocument2 pagesLe Tissu NerveuxslimdamakPas encore d'évaluation
- Tissu Nerveux - PolycopiéDocument10 pagesTissu Nerveux - PolycopiéAmina DrkPas encore d'évaluation
- Generalites IDocument11 pagesGeneralites IzqwjjbngxnPas encore d'évaluation
- 1 - Généralités Sur Le Système Nerveux 2022Document11 pages1 - Généralités Sur Le Système Nerveux 2022jack rousouPas encore d'évaluation
- Cours SN DR NaoualiDocument75 pagesCours SN DR NaoualiKobePas encore d'évaluation
- S1 Élève Organisatiion Du Système Cérébro SpinalDocument6 pagesS1 Élève Organisatiion Du Système Cérébro SpinalYacouba sanda HadizaPas encore d'évaluation
- Groupe 3Document13 pagesGroupe 3ngueguimlaurellaPas encore d'évaluation
- Tissu NerveuxDocument6 pagesTissu NerveuxOussama FendaliPas encore d'évaluation
- 5 - Physiologie Du NeuroneDocument8 pages5 - Physiologie Du Neuronemintchimechak2Pas encore d'évaluation
- Le Système NerveuxDocument7 pagesLe Système NerveuxZizou HamdiPas encore d'évaluation
- Neuro AnatDocument43 pagesNeuro AnatNouhaila AzirarPas encore d'évaluation
- La Neurophysiologie de L Appareil ManducateurDocument16 pagesLa Neurophysiologie de L Appareil ManducateurPeylost OnangelPas encore d'évaluation
- Le Déterminisme Du SNDocument25 pagesLe Déterminisme Du SNwiemPas encore d'évaluation
- Résumé G.Chakib NerveuxDocument5 pagesRésumé G.Chakib NerveuxMaroua AbzPas encore d'évaluation
- Cours SVT Tle D&C 2013-2014Document17 pagesCours SVT Tle D&C 2013-2014sandwidibernard76Pas encore d'évaluation
- Bac 2020 - Compilation Sigle en CouleurDocument76 pagesBac 2020 - Compilation Sigle en Couleurمحمد الأمين ولد عالي ابليلPas encore d'évaluation
- Documents TC SC Unit2 1Document12 pagesDocuments TC SC Unit2 1محمد الأمين ولد عالي ابليلPas encore d'évaluation
- OPATOWSKI 2020 ArchivageDocument189 pagesOPATOWSKI 2020 Archivageمحمد الأمين ولد عالي ابليلPas encore d'évaluation
- Cours Genetiques Humaine Boukhatem PDFDocument28 pagesCours Genetiques Humaine Boukhatem PDFsamabderrahimPas encore d'évaluation
- Série 3 CorrigéDocument16 pagesSérie 3 Corrigéمحمد الأمين ولد عالي ابليلPas encore d'évaluation
- TD 3 de BiochimieDocument5 pagesTD 3 de BiochimieRich HoulrichPas encore d'évaluation
- TD1+Solution Génie GénétiqueDocument5 pagesTD1+Solution Génie Génétiqueمحمد الأمين ولد عالي ابليلPas encore d'évaluation
- La Base en Bio StatiqueDocument2 pagesLa Base en Bio Statiqueمحمد الأمين ولد عالي ابليلPas encore d'évaluation
- 1 - Cours Neuropathologie Introduction Et Gliomes FMPR Anapath SpecialeDocument82 pages1 - Cours Neuropathologie Introduction Et Gliomes FMPR Anapath SpecialeMunir GuirouanPas encore d'évaluation
- Cours Séq 6 Tle D COLBICO 1Document14 pagesCours Séq 6 Tle D COLBICO 1SalifPas encore d'évaluation
- NeuroanatomieDocument113 pagesNeuroanatomiePère ProsperPas encore d'évaluation
- Physiologie Du Nerf 2022 Mede UM6Document163 pagesPhysiologie Du Nerf 2022 Mede UM6Girl Ytb100% (1)
- Cours de Neuroanatomie G2 2024Document81 pagesCours de Neuroanatomie G2 2024elohimmvulaPas encore d'évaluation
- Physio Nerv 1 FMPR PDFDocument261 pagesPhysio Nerv 1 FMPR PDFZINEB LAMRANIPas encore d'évaluation
- Organisation Du Système NerveuxDocument19 pagesOrganisation Du Système NerveuxMaelle DURANDPas encore d'évaluation
- CM Neurosciences: Le NeuroneDocument52 pagesCM Neurosciences: Le NeuroneEsther HenaultPas encore d'évaluation
- Le Tissu Nerveux - Vue GénéraleDocument67 pagesLe Tissu Nerveux - Vue GénéraleSiaka YodaPas encore d'évaluation
- Le Dictionnaire Visuel Définitions - Être HumainDocument178 pagesLe Dictionnaire Visuel Définitions - Être HumainMoncef PechaPas encore d'évaluation
- Le Programme Pour Bien Nourrir Votre Cerveau 2019 Jean Marie BourreDocument367 pagesLe Programme Pour Bien Nourrir Votre Cerveau 2019 Jean Marie BourreMoussaPas encore d'évaluation
- SYSTEME NERVEUX RechDocument14 pagesSYSTEME NERVEUX Rechariane.vasseur29Pas encore d'évaluation
- Pathologie Neurologique Périnatale Et Ses ConséquencesDocument272 pagesPathologie Neurologique Périnatale Et Ses ConséquencesAnes ArcPas encore d'évaluation
- CourDocument20 pagesCourMuhamedMouslim100% (1)
- Histologie de La Moelle ÉpinièreDocument8 pagesHistologie de La Moelle Épinièrelouanne kloepferPas encore d'évaluation
- Les Tissus NerveuxDocument9 pagesLes Tissus NerveuxZoulkiffiPas encore d'évaluation
- BIO 270 Physiologie Cellulaire Générale 2020 PDFDocument125 pagesBIO 270 Physiologie Cellulaire Générale 2020 PDFTONDI ABDOU CHACOURPas encore d'évaluation
- Banque QCM CorrigésDocument22 pagesBanque QCM CorrigésTr LwPas encore d'évaluation
- Le Cerveau en 30 Secondes PDFDocument238 pagesLe Cerveau en 30 Secondes PDFLombe Christian100% (1)
- Cerveau p17Document15 pagesCerveau p17Betschbel Dorcé100% (1)
- Lexique de Sante MentaleDocument110 pagesLexique de Sante MentaleJohn St PreuxPas encore d'évaluation
- Diapos AnatDocument142 pagesDiapos Anatjules delahayePas encore d'évaluation
- 001 Anatomie Generale Du Systeme NerveuxDocument22 pages001 Anatomie Generale Du Systeme Nerveuxشكوب ستانPas encore d'évaluation
- L 1 Neurortho 4Document34 pagesL 1 Neurortho 4fatima zahra banhssinPas encore d'évaluation
- 1 Le Systeme Nerveux I Le Tissu Nerveux Le TissuDocument13 pages1 Le Systeme Nerveux I Le Tissu Nerveux Le Tissundeye yarame diopPas encore d'évaluation
- Cours Tissus St2sDocument9 pagesCours Tissus St2sDieuboue ArthurPas encore d'évaluation
- Le Systeme NerveuxDocument22 pagesLe Systeme Nerveuxjuliette.koleczko4518Pas encore d'évaluation
- Histologie Et Biologie Cellulaire Une Introduction Áa Lanatomie PathologiqueDocument12 pagesHistologie Et Biologie Cellulaire Une Introduction Áa Lanatomie Pathologiquedjaouad bourouaisPas encore d'évaluation
- Système Nerveux 1Document21 pagesSystème Nerveux 1Ranime Mekersi100% (1)
- Neuropsychologie: Les bases théoriques et pratiques du domaine d'étude (psychologie pour tous)D'EverandNeuropsychologie: Les bases théoriques et pratiques du domaine d'étude (psychologie pour tous)Pas encore d'évaluation
- Le Pouvoir De L'Autodiscipline: L’art de tout accomplir sans limiteD'EverandLe Pouvoir De L'Autodiscipline: L’art de tout accomplir sans limiteÉvaluation : 4 sur 5 étoiles4/5 (16)
- Améliorer votre mémoire: Un Guide pour l'augmentation de la puissance du cerveau, utilisant des techniques et méthodesD'EverandAméliorer votre mémoire: Un Guide pour l'augmentation de la puissance du cerveau, utilisant des techniques et méthodesÉvaluation : 5 sur 5 étoiles5/5 (2)
- Histoire de la psychologie scientifique: De la naissance de la psychologie à la neuropsychologie et aux champs d'application les plus actuelsD'EverandHistoire de la psychologie scientifique: De la naissance de la psychologie à la neuropsychologie et aux champs d'application les plus actuelsPas encore d'évaluation
- La vie des abeilles: Prix Nobel de littératureD'EverandLa vie des abeilles: Prix Nobel de littératureÉvaluation : 4 sur 5 étoiles4/5 (41)
- Le TDAH chez l’adulte : Comment reconnaître & faire face au TDAH chez l’adulte en 30 étapes faciles.D'EverandLe TDAH chez l’adulte : Comment reconnaître & faire face au TDAH chez l’adulte en 30 étapes faciles.Pas encore d'évaluation
- Introduction à la psychologie des émotions: De Darwin aux neurosciences: découvrir les émotions et leur mode de fonctionnementD'EverandIntroduction à la psychologie des émotions: De Darwin aux neurosciences: découvrir les émotions et leur mode de fonctionnementPas encore d'évaluation
- Géologie de l'Amérique: Les Grands Articles d'UniversalisD'EverandGéologie de l'Amérique: Les Grands Articles d'UniversalisPas encore d'évaluation
- Harmonisation Energétique des Lieux: Habitat et haut-lieux sacrés 2020D'EverandHarmonisation Energétique des Lieux: Habitat et haut-lieux sacrés 2020Évaluation : 2.5 sur 5 étoiles2.5/5 (3)
- La psychologie de l'anxiété: La connaître pour comprendre ses mécanismes de fonctionnementD'EverandLa psychologie de l'anxiété: La connaître pour comprendre ses mécanismes de fonctionnementPas encore d'évaluation
- Cahier de jeux de stimulation cognitive: Sujets Alzheimer, désorientés, démences, amnésiesD'EverandCahier de jeux de stimulation cognitive: Sujets Alzheimer, désorientés, démences, amnésiesPas encore d'évaluation
- Hypnose & affirmations positives pour la richesse, l’abondance et la réussite (21 jours pour un nouveau vous)D'EverandHypnose & affirmations positives pour la richesse, l’abondance et la réussite (21 jours pour un nouveau vous)Évaluation : 5 sur 5 étoiles5/5 (1)
- La thérapie libre: La Thérapie de LIBération et Résilience EmotionnelleD'EverandLa thérapie libre: La Thérapie de LIBération et Résilience EmotionnelleÉvaluation : 5 sur 5 étoiles5/5 (1)
- La Conscience Et L'Univers Existent Sans Commencement Ni FinD'EverandLa Conscience Et L'Univers Existent Sans Commencement Ni FinPas encore d'évaluation
- Le CODE DE DIEU: Le secret de notre passé, la promesse de notre avenirD'EverandLe CODE DE DIEU: Le secret de notre passé, la promesse de notre avenirÉvaluation : 4 sur 5 étoiles4/5 (23)
- Pourquoi suis-je différent?: Accepter et découvrir la puissance de l'hypersensibilitéD'EverandPourquoi suis-je différent?: Accepter et découvrir la puissance de l'hypersensibilitéPas encore d'évaluation
- L' ESPRIT EST SON PROPRE MÉDECIN: Le pouvoir de guérison de la méditationD'EverandL' ESPRIT EST SON PROPRE MÉDECIN: Le pouvoir de guérison de la méditationPas encore d'évaluation
- Qu'arrive t'il a la terre arable ?: Le problème de l'érosion du sol.D'EverandQu'arrive t'il a la terre arable ?: Le problème de l'érosion du sol.Évaluation : 5 sur 5 étoiles5/5 (1)
- Atlas permanent de l'Union européenne: 5e édition revue et augmentéeD'EverandAtlas permanent de l'Union européenne: 5e édition revue et augmentéePas encore d'évaluation
- Connaître ses réactions émotionnelles, pour maîtriser ses émotionsD'EverandConnaître ses réactions émotionnelles, pour maîtriser ses émotionsPas encore d'évaluation
- Troubles Bipolaires: Mieux les connaître pour mieux se débarrasser de ces souffrancesD'EverandTroubles Bipolaires: Mieux les connaître pour mieux se débarrasser de ces souffrancesÉvaluation : 4.5 sur 5 étoiles4.5/5 (3)