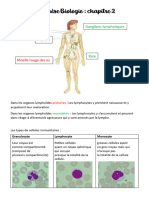
Vous aimerez peut-être aussi
- Membranes cellulaires: Les Grands Articles d'UniversalisD'EverandMembranes cellulaires: Les Grands Articles d'UniversalisPas encore d'évaluation
- Cycle de KrebsDocument36 pagesCycle de Krebskeraghel nourPas encore d'évaluation
- 3 La RéplicationDocument17 pages3 La Réplicationbahidjakheira belouadi100% (1)
- Clonage BactérienDocument35 pagesClonage Bactérienmafalada85100% (3)
- 1.introduction À L'immunologie - Résumé HS PDFDocument3 pages1.introduction À L'immunologie - Résumé HS PDFbib bib100% (1)
- ImmunologieDocument13 pagesImmunologieXzistence100% (10)
- Déroulement de La Réponse Immunitaire SpécifiqueDocument6 pagesDéroulement de La Réponse Immunitaire SpécifiqueDalili SVTPas encore d'évaluation
- Le Systeme ImmunitaireDocument32 pagesLe Systeme ImmunitaireIlyes MraidiaPas encore d'évaluation
- 05 Les Molecules D - AdhesionDocument4 pages05 Les Molecules D - AdhesionOns H'maiedPas encore d'évaluation
- Cour4 Expression D'antigènes, Marqueurs de Virulence Et de Récepteurs CellulairesDocument6 pagesCour4 Expression D'antigènes, Marqueurs de Virulence Et de Récepteurs CellulairesJoie De VivrePas encore d'évaluation
- Cour3 Les Molécules Dadhésion CellulaireDocument3 pagesCour3 Les Molécules Dadhésion CellulaireJoie De VivrePas encore d'évaluation
- Molécules D'adhérence Cours L1Document3 pagesMolécules D'adhérence Cours L1safemindPas encore d'évaluation
- Cours de Biologie Cellulaire L3 BMCDocument16 pagesCours de Biologie Cellulaire L3 BMCIlyasse AitPas encore d'évaluation
- I07. Cytokines (Cour)Document7 pagesI07. Cytokines (Cour)Hibatallah NamounePas encore d'évaluation
- Résumé GCDocument3 pagesRésumé GCLilia TlbPas encore d'évaluation
- Les Cytokines 2019 2020Document7 pagesLes Cytokines 2019 2020tara mouradPas encore d'évaluation
- Cours #1 - Jonction CellDocument96 pagesCours #1 - Jonction CellMzn JannahPas encore d'évaluation
- Biologie CellulaireDocument297 pagesBiologie Cellulaireesmorad53Pas encore d'évaluation
- Adherence 2017Document26 pagesAdherence 2017laarabPas encore d'évaluation
- Chap 3 Les Protéines Impliquées Dans La MECDocument7 pagesChap 3 Les Protéines Impliquées Dans La MECLOLOPas encore d'évaluation
- Biologie Générale HumaineDocument10 pagesBiologie Générale HumaineSarah CabieuPas encore d'évaluation
- BIO111 ChapitreI Suite 01Document5 pagesBIO111 ChapitreI Suite 01Berthe Aristide Ngue Nkongo100% (1)
- CytologieDocument7 pagesCytologiedorissePas encore d'évaluation
- Sujeujet Bio Celle Session 2 PCEM1Document4 pagesSujeujet Bio Celle Session 2 PCEM1medPas encore d'évaluation
- Chapitre II Membrane Plasmique 1Document7 pagesChapitre II Membrane Plasmique 1faresgettasPas encore d'évaluation
- ImmunoDocument5 pagesImmunoAmadou CISSEPas encore d'évaluation
- ChondriomeDocument25 pagesChondriomeYlinka SamiraPas encore d'évaluation
- Biologie - Cellulaire - Cour KamelDocument15 pagesBiologie - Cellulaire - Cour Kamelrekik hibaPas encore d'évaluation
- UE2 - Biologie Cellulaire - Molécule de Surface Et D - AdhérenceDocument19 pagesUE2 - Biologie Cellulaire - Molécule de Surface Et D - Adhérencesun-nee-chan9Pas encore d'évaluation
- Chap - 3 - Les Molécules de SurfaceDocument6 pagesChap - 3 - Les Molécules de SurfaceMoeanau GrangerPas encore d'évaluation
- Synthèse Biologie chp2Document6 pagesSynthèse Biologie chp2clara.plv22Pas encore d'évaluation
- Cours BCG UFR2SDocument19 pagesCours BCG UFR2SSirandou DiaoPas encore d'évaluation
- Biologie CellulaireDocument26 pagesBiologie CellulaireBourobou bourobou Chadrack gaelPas encore d'évaluation
- Chapitre 2Document7 pagesChapitre 2dyhp2pswsrPas encore d'évaluation
- Cours MECDocument14 pagesCours MEChabenu100% (1)
- Epreuve Bio Cell - Numerus - 2021-2022Document3 pagesEpreuve Bio Cell - Numerus - 2021-2022Mohamed DickoPas encore d'évaluation
- La Réponse Immunitaire Innée Et Adaptative ModifDocument8 pagesLa Réponse Immunitaire Innée Et Adaptative ModifAni TrghwiPas encore d'évaluation
- Cours Immunologie - 2ème Sciences Biologiques Section 01 - OUABEDDocument22 pagesCours Immunologie - 2ème Sciences Biologiques Section 01 - OUABEDKakashi HatakéPas encore d'évaluation
- Chap - 2 - Les Phénomènes de TransportDocument5 pagesChap - 2 - Les Phénomènes de TransportMoeanau GrangerPas encore d'évaluation
- Notes Bio CellDocument14 pagesNotes Bio CellYoussef LahjomriPas encore d'évaluation
- Cours Histologie 2 Partie 2Document66 pagesCours Histologie 2 Partie 2Salomon JosephPas encore d'évaluation
- Httpselearning Centre-Univ-Mila Dza2024pluginfile Php89198mod - resourcecontent1CHAPITRE20V PDFDocument20 pagesHttpselearning Centre-Univ-Mila Dza2024pluginfile Php89198mod - resourcecontent1CHAPITRE20V PDFariel.2mariemPas encore d'évaluation
- Cours Immunologie - 2ème Sciences Biologiques Section 01 - OUABEDDocument19 pagesCours Immunologie - 2ème Sciences Biologiques Section 01 - OUABEDDjihadnoorPas encore d'évaluation
- Sheets Du Dossier GpoiesefDocument7 pagesSheets Du Dossier Gpoiesefyaya camaraPas encore d'évaluation
- Croissance CellulaireDocument28 pagesCroissance Cellulaireهوبة سفيلPas encore d'évaluation
- Fiche CMHDocument3 pagesFiche CMHrequiemforathrowawayPas encore d'évaluation
- RappelDocument11 pagesRappelNadjib A. NedjarPas encore d'évaluation
- 2022 MTC - DR MeradDocument8 pages2022 MTC - DR Meradkhouloudnemmouchi240721Pas encore d'évaluation
- UE Endocrinologie Cellulaire M2PANDocument13 pagesUE Endocrinologie Cellulaire M2PANteresa ekouerembaPas encore d'évaluation
- BIOCELL3Document89 pagesBIOCELL3samPas encore d'évaluation
- Immunologie - Cahier PRDocument64 pagesImmunologie - Cahier PRJulie LEMOINEPas encore d'évaluation
- C M 2-Oncologie I-2016Document111 pagesC M 2-Oncologie I-2016NanfaPas encore d'évaluation
- La Matrice Extracellulaire 2015-2016 PDFDocument3 pagesLa Matrice Extracellulaire 2015-2016 PDFAnonymous wVr908lBcK100% (2)
- Solution Des Exercices Le Système ImmunitaireDocument15 pagesSolution Des Exercices Le Système ImmunitairendiayePas encore d'évaluation
- Membrane PlasmiqueDocument17 pagesMembrane PlasmiqueSami SamirPas encore d'évaluation
- Chap 2 MPDocument7 pagesChap 2 MPdramohandlhadjPas encore d'évaluation
- 3ème Partie Du Chapitre 1 Membrane Plasmique La Communication CellulaireDocument46 pages3ème Partie Du Chapitre 1 Membrane Plasmique La Communication CellulaireElHamdouni ZiadPas encore d'évaluation
- Membrane PlasmiqueDocument7 pagesMembrane PlasmiqueSara BoudjemaiPas encore d'évaluation
- Fichier Produit 3806Document2 pagesFichier Produit 3806BOULAMA HASSANA MASSAWPas encore d'évaluation
- BIO 111 - CHAP - VI - VII - Et - VIIIDocument9 pagesBIO 111 - CHAP - VI - VII - Et - VIIIBerthe Aristide Ngue NkongoPas encore d'évaluation
- 1.cours Biologie CellulaireDocument63 pages1.cours Biologie Cellulaireبدر الذهابيPas encore d'évaluation
- Httpsstaff - Univ-Batna2.dzsitesdefaultfilesabdessemed Samirafileschapitre 11 PDFDocument12 pagesHttpsstaff - Univ-Batna2.dzsitesdefaultfilesabdessemed Samirafileschapitre 11 PDFariel.2mariemPas encore d'évaluation
- I02. Cellules de L'immunité (Cour)Document24 pagesI02. Cellules de L'immunité (Cour)Uubay 2003Pas encore d'évaluation
- TP ADNDocument3 pagesTP ADNNaomi LAMBERTPas encore d'évaluation
- 2014 03 NelleCaledo Exo3 Sujet Peptide Protection 5ptsDocument3 pages2014 03 NelleCaledo Exo3 Sujet Peptide Protection 5ptsHaman Bello OumarouPas encore d'évaluation
- Chimiothérapie AntiviraleDocument25 pagesChimiothérapie AntiviraleKawtar Kaaboub67% (3)
- Biochimie Metabolique Les Glucides CoursDocument20 pagesBiochimie Metabolique Les Glucides CoursmaryPas encore d'évaluation
- AntibiotiquesDocument15 pagesAntibiotiquesbelarouci linaPas encore d'évaluation
- Le Génie Génétique AgroDocument112 pagesLe Génie Génétique AgroWa LidPas encore d'évaluation
- Devoir de Contrôle N°2 - SVT - 3ème SC Exp (2009-2010)Document3 pagesDevoir de Contrôle N°2 - SVT - 3ème SC Exp (2009-2010)sonilola100% (1)
- Introduction Au MétabolismeDocument8 pagesIntroduction Au MétabolismeAtayiPas encore d'évaluation
- Partie II - Bio Cellulaire - II.A.Les Procaryotes - 5. Synthèse Des ProtéinesDocument83 pagesPartie II - Bio Cellulaire - II.A.Les Procaryotes - 5. Synthèse Des ProtéinesmanelbeggarPas encore d'évaluation
- Vecteurs de ClonageDocument37 pagesVecteurs de Clonagelobna75% (4)
- TD N°1 en Biologie Moléculaire Et Génie GénétiqueDocument4 pagesTD N°1 en Biologie Moléculaire Et Génie GénétiqueBemmoussat AsmaPas encore d'évaluation
- TSTL - Bioch.Cours .Chap.2.Enzymologie - Correction Exercice.7Document3 pagesTSTL - Bioch.Cours .Chap.2.Enzymologie - Correction Exercice.7Chris Zericho0% (3)
- Virologie Virologie.Document8 pagesVirologie Virologie.Sarra FerroudjiPas encore d'évaluation
- UdeM Maîtrise en Biologie Moléculaire-2022-07!24!130356Document7 pagesUdeM Maîtrise en Biologie Moléculaire-2022-07!24!130356tankarirouwanePas encore d'évaluation
- Corrige-Type Travail Personnel Genie Genetique 3eme Annee LMD BiochimieDocument2 pagesCorrige-Type Travail Personnel Genie Genetique 3eme Annee LMD Biochimieamina44Pas encore d'évaluation
- Biologie Cellulaire Medecine - CopieDocument152 pagesBiologie Cellulaire Medecine - CopieFulbert KiemaPas encore d'évaluation
- Biologie Moléculaire Des Plantes - 8 NovembreDocument16 pagesBiologie Moléculaire Des Plantes - 8 Novembrezuzusmail12Pas encore d'évaluation
- Noyau Mitoch PeroxDocument138 pagesNoyau Mitoch PeroxHamza HadoutiPas encore d'évaluation
- Notes Bio CellDocument14 pagesNotes Bio CellYoussef LahjomriPas encore d'évaluation
- Techniques de Quantification de La CVP VIH-1 VVDocument45 pagesTechniques de Quantification de La CVP VIH-1 VVAmadou DraboPas encore d'évaluation
- La Paroi BacterienneDocument40 pagesLa Paroi BacterienneBile DjetouanPas encore d'évaluation
- Sdic PL0916Document45 pagesSdic PL0916Ahmed DoghmiPas encore d'évaluation
- C SVTDocument3 pagesC SVTAyn RATSIMBAZAFYPas encore d'évaluation
- PDF CHAPITRE No2 La Variabilite Genetique Et MutationsDocument15 pagesPDF CHAPITRE No2 La Variabilite Genetique Et MutationssimplicitaPas encore d'évaluation
- TD1 EnzymologieDocument7 pagesTD1 EnzymologieasmaPas encore d'évaluation
- Fiche Méthode - Synthèse PeptidiqueDocument2 pagesFiche Méthode - Synthèse PeptidiqueTaleen PekerPas encore d'évaluation
- Chapitre1 Cycle CellulaireDocument17 pagesChapitre1 Cycle CellulaireEnnassiki Med EnnassikiPas encore d'évaluation