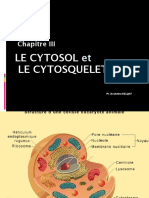
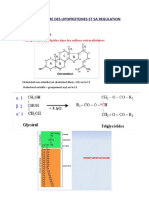
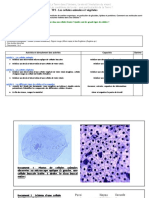
Vous aimerez peut-être aussi
- BIOCELL3Document89 pagesBIOCELL3samPas encore d'évaluation
- UE2 Le CytosqueletteDocument18 pagesUE2 Le Cytosquelettesun-nee-chan9Pas encore d'évaluation
- La Matrice Extracellulaire 2015-2016 PDFDocument3 pagesLa Matrice Extracellulaire 2015-2016 PDFAnonymous wVr908lBcK100% (2)
- Chapitre 3. Hyaloplasme Et Cytosquelette 1Document6 pagesChapitre 3. Hyaloplasme Et Cytosquelette 1Yahia BerbouchiPas encore d'évaluation
- BiocellPCEM12002 2003 Cours MaJ 2003 04 11Document58 pagesBiocellPCEM12002 2003 Cours MaJ 2003 04 11Gilbert ThierryPas encore d'évaluation
- Le Cytosquelette - PpsDocument14 pagesLe Cytosquelette - PpsnasredinnePas encore d'évaluation
- Les Micro FilamentsDocument8 pagesLes Micro FilamentsYouness KhalfaouiPas encore d'évaluation
- Le CytoplasmeDocument15 pagesLe CytoplasmeWissam TizaPas encore d'évaluation
- CytosqueletteDocument27 pagesCytosqueletteFatima BENHAFSSAPas encore d'évaluation
- Le Cytosquelette. 2021 CH7Document68 pagesLe Cytosquelette. 2021 CH7paul destinPas encore d'évaluation
- Cours de Biologie Cellulaire L3 BMCDocument16 pagesCours de Biologie Cellulaire L3 BMCIlyasse AitPas encore d'évaluation
- Chapitre 7Document10 pagesChapitre 7bousbibitsaraPas encore d'évaluation
- Chapitre 2Document7 pagesChapitre 2dyhp2pswsrPas encore d'évaluation
- Cours BCG UFR2SDocument19 pagesCours BCG UFR2SSirandou DiaoPas encore d'évaluation
- CytosqueletteDocument3 pagesCytosqueletteFares BENHENDAPas encore d'évaluation
- Le Tissu Conjonctif Non Spécialisé WordDocument10 pagesLe Tissu Conjonctif Non Spécialisé WordAbdoulaziz SidilaminePas encore d'évaluation
- Structure Et Physiologie Des Eucaryotes 2018-2019-L2S4Document7 pagesStructure Et Physiologie Des Eucaryotes 2018-2019-L2S4Original ZeyePas encore d'évaluation
- Le CytosqueletteDocument9 pagesLe CytosqueletteAmï NãPas encore d'évaluation
- Chap - 4 - Les Jonctions IntercellulairesDocument5 pagesChap - 4 - Les Jonctions IntercellulairesMoeanau GrangerPas encore d'évaluation
- 1 Cours de Biologie CellulaireDocument18 pages1 Cours de Biologie CellulaireSamy Ta'y'b'iPas encore d'évaluation
- Généralités Sur La CelluleDocument14 pagesGénéralités Sur La CelluleJulien MinotPas encore d'évaluation
- TD 3 Le CytosqueletteDocument7 pagesTD 3 Le CytosqueletteĐî ŅæPas encore d'évaluation
- Résumé GCDocument3 pagesRésumé GCLilia TlbPas encore d'évaluation
- Biologie CellulaireDocument297 pagesBiologie Cellulaireesmorad53Pas encore d'évaluation
- CytosqueletteDocument17 pagesCytosqueletteKa TiaPas encore d'évaluation
- Fichier Produit 3806Document2 pagesFichier Produit 3806BOULAMA HASSANA MASSAWPas encore d'évaluation
- Cours MECDocument14 pagesCours MEChabenu100% (1)
- Structure Des Bactéries 2018 2019Document8 pagesStructure Des Bactéries 2018 2019Ismail HamrouniPas encore d'évaluation
- Quelle Est L'ultrastructure de La Cellule Eucaryote ?Document5 pagesQuelle Est L'ultrastructure de La Cellule Eucaryote ?youseftriki7Pas encore d'évaluation
- HyaloplasmeDocument25 pagesHyaloplasmeamina44Pas encore d'évaluation
- Reste Du Chapitre CytosqueletteDocument6 pagesReste Du Chapitre CytosqueletteMohammed El MachmoulPas encore d'évaluation
- Cours 1 CytosqueletteDocument12 pagesCours 1 CytosquelettenadeauPas encore d'évaluation
- Le Cytosquelette1Document18 pagesLe Cytosquelette1Radjaa BerroubaPas encore d'évaluation
- Le CytoplasmeDocument13 pagesLe CytoplasmeAIFRAPas encore d'évaluation
- Cours de Cytologie, Embryologie Et Genetique Medicale by MBAILASSEM MBAINAIDA GEORGESDocument102 pagesCours de Cytologie, Embryologie Et Genetique Medicale by MBAILASSEM MBAINAIDA GEORGESMãhąmãţ Hãbįb Mõųşşã ĄhmąţPas encore d'évaluation
- Exposé Biochimie Cellulaire Et FonctionnelleDocument12 pagesExposé Biochimie Cellulaire Et FonctionnelleairaPas encore d'évaluation
- CH3 - Cytosol-Cytosquelette - B.BDocument47 pagesCH3 - Cytosol-Cytosquelette - B.Bone asslanPas encore d'évaluation
- Capture D'écran . 2022-04-22 À 23.46.45Document46 pagesCapture D'écran . 2022-04-22 À 23.46.45Lana SeaPas encore d'évaluation
- Chapitre 1 Histologie-ConvertiDocument10 pagesChapitre 1 Histologie-ConvertiMond DayPas encore d'évaluation
- Correction - TD2 - BI - 125 - Structure - Et Fonction - de La - Cellule - AnimaleDocument5 pagesCorrection - TD2 - BI - 125 - Structure - Et Fonction - de La - Cellule - AnimaleBerthe Aristide Ngue NkongoPas encore d'évaluation
- Cytosquelette 1Document13 pagesCytosquelette 1Nadine DelormePas encore d'évaluation
- Matrice BPC 2Document39 pagesMatrice BPC 2Mzn JannahPas encore d'évaluation
- Chap 01Document33 pagesChap 01hamidaPas encore d'évaluation
- Master 2 (Chap.1)Document6 pagesMaster 2 (Chap.1)Ažį ŽaPas encore d'évaluation
- Chap 2 MPDocument7 pagesChap 2 MPdramohandlhadjPas encore d'évaluation
- Chapitre 2 BiomembraneDocument5 pagesChapitre 2 BiomembraneFatïma ZohraPas encore d'évaluation
- Epreuve Bio Cell - Numerus - 2021-2022Document3 pagesEpreuve Bio Cell - Numerus - 2021-2022Mohamed DickoPas encore d'évaluation
- Membrane PlasmiqueDocument7 pagesMembrane PlasmiqueSara BoudjemaiPas encore d'évaluation
- La Membrane Plasmique: I. GénéralitésDocument17 pagesLa Membrane Plasmique: I. GénéralitésAisSa KeRrøùùmPas encore d'évaluation
- Chapitre III Cours CytosqueletteDocument4 pagesChapitre III Cours CytosquelettefaresgettasPas encore d'évaluation
- Chapitre Xii Le Cytosquelette PDFDocument17 pagesChapitre Xii Le Cytosquelette PDFRaniaPas encore d'évaluation
- PosterDocument5 pagesPosterBrahim OuarhlentPas encore d'évaluation
- Cours Annuel SVT EDUCCIADocument96 pagesCours Annuel SVT EDUCCIAYvanna TebeuPas encore d'évaluation
- Résumé 3Document4 pagesRésumé 3Farah BaaraPas encore d'évaluation
- Adherence 2017Document26 pagesAdherence 2017laarabPas encore d'évaluation
- Chap 3 MEC 11Document16 pagesChap 3 MEC 11Sara BoudjemaiPas encore d'évaluation
- Membrane PlasmiqueDocument17 pagesMembrane PlasmiqueSami SamirPas encore d'évaluation
- Membranes cellulaires: Les Grands Articles d'UniversalisD'EverandMembranes cellulaires: Les Grands Articles d'UniversalisPas encore d'évaluation
- Champignons: Les Grands Articles d'UniversalisD'EverandChampignons: Les Grands Articles d'UniversalisPas encore d'évaluation
- Muscles: Les Grands Articles d'UniversalisD'EverandMuscles: Les Grands Articles d'UniversalisPas encore d'évaluation
- TD Génomique Fonctionnelle 2022 - 2023Document96 pagesTD Génomique Fonctionnelle 2022 - 2023rekik hibaPas encore d'évaluation
- Fichier Produit 3082Document3 pagesFichier Produit 3082rekik hibaPas encore d'évaluation
- Pdf2star 1476439438 Manuscrit TH Se AG Sicaire FINALDocument227 pagesPdf2star 1476439438 Manuscrit TH Se AG Sicaire FINALHardy BONDAPas encore d'évaluation
- Lextraction Des Composées NaturellesDocument6 pagesLextraction Des Composées Naturellesrekik hibaPas encore d'évaluation
- NEBEG HalimaDocument136 pagesNEBEG Halimarekik hibaPas encore d'évaluation
- (123dok - Net) Etude de L Activité Antioxydante Des Saponines de L ArganierDocument149 pages(123dok - Net) Etude de L Activité Antioxydante Des Saponines de L Arganierrekik hibaPas encore d'évaluation
- 2013 Actualit ChimiqueDocument6 pages2013 Actualit Chimiquerekik hibaPas encore d'évaluation
- Polycopie ThermoDocument116 pagesPolycopie Thermorekik hibaPas encore d'évaluation
- ThèseDocument188 pagesThèseAli AsdadiPas encore d'évaluation
- Chimie Generala Corection Serie 1Document4 pagesChimie Generala Corection Serie 1rekik hibaPas encore d'évaluation
- CJ-VieQuotidienne 8Document15 pagesCJ-VieQuotidienne 8rekik hibaPas encore d'évaluation
- Cloutier Uqac 0862N 11054Document66 pagesCloutier Uqac 0862N 11054rekik hibaPas encore d'évaluation
- Examan BADocument4 pagesExaman BArekik hibaPas encore d'évaluation
- Electricite DomestiqueDocument15 pagesElectricite Domestiqueyabapo5957Pas encore d'évaluation
- Série 1 Complément MathDocument4 pagesSérie 1 Complément Mathrekik hibaPas encore d'évaluation
- Série 1 Chimie Generale AU 2020Document1 pageSérie 1 Chimie Generale AU 2020rekik hibaPas encore d'évaluation
- Math TDDocument6 pagesMath TDrekik hibaPas encore d'évaluation
- Cette StratégieDocument2 pagesCette Stratégierekik hibaPas encore d'évaluation
- Fascicule TP BA2 2019 2020Document58 pagesFascicule TP BA2 2019 2020rekik hiba100% (2)
- Genetique TDDocument4 pagesGenetique TDrekik hibaPas encore d'évaluation
- Embryologie.2020-2021.partie 2 (Mode de Compatibilité)Document19 pagesEmbryologie.2020-2021.partie 2 (Mode de Compatibilité)rekik hibaPas encore d'évaluation
- ManipDocument4 pagesManiprekik hibaPas encore d'évaluation
- Metabolisme Des Lipoproteines Et Sa RegulationDocument8 pagesMetabolisme Des Lipoproteines Et Sa Regulationrekik hibaPas encore d'évaluation
- I. But:: Rekik Hiba Tp2: Influence de PH Sur L'activité de L'invertaseDocument3 pagesI. But:: Rekik Hiba Tp2: Influence de PH Sur L'activité de L'invertaserekik hibaPas encore d'évaluation
- Chez La TomateDocument2 pagesChez La Tomaterekik hibaPas encore d'évaluation
- Chapitre 3 Maturation Des ProtéinesDocument34 pagesChapitre 3 Maturation Des Protéinesryadh2203Pas encore d'évaluation
- Les CytokinesDocument47 pagesLes CytokinesOusmane SyllaPas encore d'évaluation
- Cours 2Document39 pagesCours 2fatiPas encore d'évaluation
- TD7 Mécanismes-Moléculaires-Respiration Correction PDFDocument3 pagesTD7 Mécanismes-Moléculaires-Respiration Correction PDFGabin Dossou100% (1)
- Devoir Corrige de Synthese N3 SVT 2eme SDocument8 pagesDevoir Corrige de Synthese N3 SVT 2eme Schafitalbi72Pas encore d'évaluation
- PDF NeigeDocument3 pagesPDF NeigeZineb ELPas encore d'évaluation
- h04 Trimoreau Accreditation - NfsDocument84 pagesh04 Trimoreau Accreditation - NfsMichael MouffoPas encore d'évaluation
- Bioch Pharm2an-Metabolisme GlucidesDocument28 pagesBioch Pharm2an-Metabolisme GlucidesmaryPas encore d'évaluation
- EchangesDocument12 pagesEchangesAmina assala Hadj MohamedPas encore d'évaluation
- Les Feuilles Des Monocotylédones Et Des DicotylédonesDocument2 pagesLes Feuilles Des Monocotylédones Et Des DicotylédonesMoussaba DoumbouyaPas encore d'évaluation
- TP3 Les CellulesDocument6 pagesTP3 Les CellulestaragadabbPas encore d'évaluation
- Livre Du Prof 2nde C 1Document74 pagesLivre Du Prof 2nde C 1Kadjo AnvoPas encore d'évaluation
- TD1 Programmation Cellulaire - CorrigéDocument5 pagesTD1 Programmation Cellulaire - Corrigéad5271439Pas encore d'évaluation
- Synthese Des ProteinesDocument2 pagesSynthese Des Proteinesdiawbachir15Pas encore d'évaluation
- 03 Cellule CancéreuseDocument8 pages03 Cellule CancéreuseYoucef KeddarPas encore d'évaluation
- Cours Proteines L2 SVT FST 2022-23Document28 pagesCours Proteines L2 SVT FST 2022-23Kaba NiambelePas encore d'évaluation
- PDF QCM1 Métabolisme-GlucidiqueDocument1 pagePDF QCM1 Métabolisme-GlucidiqueYendouli LAMBONIPas encore d'évaluation
- Diufral QCM Module0 Immunologie Corrige 2014 2Document4 pagesDiufral QCM Module0 Immunologie Corrige 2014 2olgaj mouadoPas encore d'évaluation
- 9-Generalites Sur Les HormonesDocument5 pages9-Generalites Sur Les HormonesoutsiderPas encore d'évaluation
- COURS - BIO 122 - Microbiologie Générale-1Document55 pagesCOURS - BIO 122 - Microbiologie Générale-1TONDI ABDOU CHACOURPas encore d'évaluation
- SVT - Bacc - Enoncé Science de La Vie Et de La Terre Série C 2004Document3 pagesSVT - Bacc - Enoncé Science de La Vie Et de La Terre Série C 2004Setra FinaritraPas encore d'évaluation
- LYMPHOPOIESEDocument5 pagesLYMPHOPOIESEyaya camara100% (1)
- Biologie CellulaireDocument25 pagesBiologie CellulaireSousou SalimPas encore d'évaluation
- Arguments Pour La GenetiqueDocument3 pagesArguments Pour La Genetiquemennarais55Pas encore d'évaluation
- Exposé Du Groupe 9Document35 pagesExposé Du Groupe 9Grâce KABOREPas encore d'évaluation
- Td4 Structure de La MP 2019-2020Document2 pagesTd4 Structure de La MP 2019-2020Yugen UniquePas encore d'évaluation
- L'INFLAMMATIO'NDocument37 pagesL'INFLAMMATIO'NLoly LolyPas encore d'évaluation
- MethylationDocument11 pagesMethylationAdli OthmanePas encore d'évaluation
- Devoir de Synthèse N°2 - SVT - 2ème Sciences Exp (2016-2017) MR Hakim NAOUELDocument2 pagesDevoir de Synthèse N°2 - SVT - 2ème Sciences Exp (2016-2017) MR Hakim NAOUELaliPas encore d'évaluation
- BiologieDocument712 pagesBiologieZakaria Wifmed100% (9)