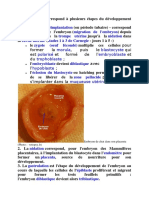
Vous aimerez peut-être aussi
- PharmacologieDocument140 pagesPharmacologievinmegaPas encore d'évaluation
- Embryologie Descriptive Chez Les OiseauxDocument10 pagesEmbryologie Descriptive Chez Les OiseauxGASMI ABDELNOURPas encore d'évaluation
- EmbryologieDocument14 pagesEmbryologiedanica2009Pas encore d'évaluation
- Deuxieme SemaineDocument5 pagesDeuxieme SemaineLysongo OruPas encore d'évaluation
- Développement Embryonaire HumaineDocument11 pagesDéveloppement Embryonaire HumaineWijdene SassiPas encore d'évaluation
- 6-Première Semaine Du Développement EmbryonnaireDocument2 pages6-Première Semaine Du Développement Embryonnaire130660LYC OMAR BENABDELAZIZ NEDROMAPas encore d'évaluation
- DVT EmbryonnaireDocument7 pagesDVT Embryonnairedyhp2pswsrPas encore d'évaluation
- Embryo TPDocument18 pagesEmbryo TPsmaalia oumaimaPas encore d'évaluation
- 2 La Deuxième Semaine Cours Final Etudiants 2023Document7 pages2 La Deuxième Semaine Cours Final Etudiants 2023rolemodel663Pas encore d'évaluation
- 1 Premiere Semaine Final Cours Etudiants 2023Document4 pages1 Premiere Semaine Final Cours Etudiants 2023rolemodel663Pas encore d'évaluation
- Développement EmbryonnaireDocument9 pagesDéveloppement EmbryonnaireEva Peignier100% (1)
- Cours 5 Segmentation Format 2Document3 pagesCours 5 Segmentation Format 2Nãřímäñ KŁayPas encore d'évaluation
- I. DéfinitionDocument2 pagesI. Définitionhammou HoudjedjePas encore d'évaluation
- Microsoft Word - Suite Chapitre IIIDocument21 pagesMicrosoft Word - Suite Chapitre IIImanarhajji2024Pas encore d'évaluation
- Develepomment Embryonnaire UsthbDocument22 pagesDevelepomment Embryonnaire UsthbMahieddine BoucekkinePas encore d'évaluation
- 647m1 EMBRYOLOGIEDocument20 pages647m1 EMBRYOLOGIEKamilia BilalPas encore d'évaluation
- Aspect Cellulaire de DéveloppementDocument9 pagesAspect Cellulaire de DéveloppementOm Nawfal NawfalPas encore d'évaluation
- (Former La, Puis Le Qui Est Et Formé de L' Et Du Avec L' Et L' Pour Pouvoir S'implanter Dans LaDocument2 pages(Former La, Puis Le Qui Est Et Formé de L' Et Du Avec L' Et L' Pour Pouvoir S'implanter Dans LaRihem BarketPas encore d'évaluation
- Texte 2e SemaineDocument7 pagesTexte 2e Semainej29024688Pas encore d'évaluation
- Embryologie Humaine GénéraleDocument13 pagesEmbryologie Humaine GénéraleHugues Roodly JeannitePas encore d'évaluation
- La SegmentationDocument18 pagesLa SegmentationNabil holmesPas encore d'évaluation
- Développement Placentaire - Cours Maïeutique JOGUET P1 03-2011 - UE8 - UE11Document4 pagesDéveloppement Placentaire - Cours Maïeutique JOGUET P1 03-2011 - UE8 - UE11Université des AntillesPas encore d'évaluation
- Embryogenèse HumaineDocument46 pagesEmbryogenèse HumaineAmbdoulwakil MohamedPas encore d'évaluation
- 1eì Re Semaine de DeveloppementDocument31 pages1eì Re Semaine de DeveloppementMøüštãphå MëdPas encore d'évaluation
- Developpement EmbryonnaireDocument18 pagesDeveloppement Embryonnaireberkatimanel22Pas encore d'évaluation
- Deuxieme Semaine Du Developpement Humain L1S2 21 Mai 2021Document30 pagesDeuxieme Semaine Du Developpement Humain L1S2 21 Mai 2021nanemafabricewilliamPas encore d'évaluation
- Chapitre V Gastrulation (Cours)Document2 pagesChapitre V Gastrulation (Cours)jsaadoudanbakoPas encore d'évaluation
- Chapitre 5 NIDATION ET PREGASTRULATIONDocument6 pagesChapitre 5 NIDATION ET PREGASTRULATIONVéto SamPas encore d'évaluation
- EmbriogeneseDocument16 pagesEmbriogeneseYahiaab RahmaniPas encore d'évaluation
- Deuxième Semaine DR EZINAHDocument33 pagesDeuxième Semaine DR EZINAHClovis Zue EssandonePas encore d'évaluation
- La 4eme-8eme Semaine Cours 2018Document4 pagesLa 4eme-8eme Semaine Cours 2018130660LYC OMAR BENABDELAZIZ NEDROMAPas encore d'évaluation
- BIOLOGIE DU DEVELOPPEMENT-La GastrulationDocument9 pagesBIOLOGIE DU DEVELOPPEMENT-La GastrulationYves DoumingouPas encore d'évaluation
- Embryologie Pour OrthodontistesDocument44 pagesEmbryologie Pour OrthodontistesHaydar AL Hasan100% (1)
- Le Disque Embryonnaire DidermiqueDocument22 pagesLe Disque Embryonnaire DidermiqueNã DāPas encore d'évaluation
- Chap 4Document13 pagesChap 4Mamadou lamine DiattaPas encore d'évaluation
- EmbryogénèseDocument5 pagesEmbryogénèseredPas encore d'évaluation
- 8 - PlacentaDocument12 pages8 - Placentalakeabdellah122.mPas encore d'évaluation
- Embryo23 05-2mois Devellopement PDFDocument30 pagesEmbryo23 05-2mois Devellopement PDFChouaib MeraoumiaPas encore d'évaluation
- 1-Anatomie Et Physiologie ObstetricalesDocument49 pages1-Anatomie Et Physiologie Obstetricalesremi degleyPas encore d'évaluation
- GastrulationDocument5 pagesGastrulationMinaheta Guy maurelPas encore d'évaluation
- Embryogenèse Et Developpement Foetal 2024Document14 pagesEmbryogenèse Et Developpement Foetal 2024KobePas encore d'évaluation
- 7-Deuxième Semaine de Développement EmbryonnaireDocument3 pages7-Deuxième Semaine de Développement Embryonnaire130660LYC OMAR BENABDELAZIZ NEDROMAPas encore d'évaluation
- 6 GastrulationDocument27 pages6 GastrulationCó TênPas encore d'évaluation
- Annexes Du FoetusDocument16 pagesAnnexes Du FoetusAMI ChanPas encore d'évaluation
- La Segmentation Chez L'oiseauDocument11 pagesLa Segmentation Chez L'oiseauUMagis Itsfx-Itsa AntadyPas encore d'évaluation
- Première Semaine EmbryologieDocument5 pagesPremière Semaine EmbryologiediayeloupPas encore d'évaluation
- La Troisième Semaine Du Dével OKDocument8 pagesLa Troisième Semaine Du Dével OKEdna windiam LaubouetPas encore d'évaluation
- Embryologie - Mise en Place de L'appareil Uro-GénitalDocument6 pagesEmbryologie - Mise en Place de L'appareil Uro-GénitalLouis PoddevinPas encore d'évaluation
- La Gametogenese CoursDocument13 pagesLa Gametogenese Coursقادة قناية100% (4)
- Embryologie.1ème Semaine de DEDocument28 pagesEmbryologie.1ème Semaine de DEHoussem MlayehPas encore d'évaluation
- 1ère Semaine OkDocument56 pages1ère Semaine OkAÏCHATA KonéPas encore d'évaluation
- 1er Semaine de Developpement EmbrynaireDocument3 pages1er Semaine de Developpement EmbrynaireHadjer TalbiPas encore d'évaluation
- L Ovogenese PDFDocument18 pagesL Ovogenese PDFSahar Mekki100% (1)
- A. Copie de 4ème Semaine deDocument3 pagesA. Copie de 4ème Semaine deILHEM RAMDA100% (1)
- ExtraitspeSVT1ere PDFDocument9 pagesExtraitspeSVT1ere PDFSafa OJEILPas encore d'évaluation
- PR Sahraoui La GestationDocument3 pagesPR Sahraoui La Gestation4cprqpbhw5Pas encore d'évaluation
- La Première Semaine de DeveloppementDocument56 pagesLa Première Semaine de DeveloppementFORN01 Dams01Pas encore d'évaluation
- Cours 4 Deuxième Semaine Du Développement Embryonnaire Nov 2023Document8 pagesCours 4 Deuxième Semaine Du Développement Embryonnaire Nov 2023madimalak111Pas encore d'évaluation
- Biologie Cellulaire IIIDocument12 pagesBiologie Cellulaire IIIInas HassanatPas encore d'évaluation
- Champignons: Les Grands Articles d'UniversalisD'EverandChampignons: Les Grands Articles d'UniversalisPas encore d'évaluation
- Développement végétal: Les Grands Articles d'UniversalisD'EverandDéveloppement végétal: Les Grands Articles d'UniversalisPas encore d'évaluation
- Quatrième Semaine Du Développement EmbryonnaireDocument9 pagesQuatrième Semaine Du Développement EmbryonnaireDrissPas encore d'évaluation
- 07 Morpho Ga StrulDocument6 pages07 Morpho Ga Strulan warPas encore d'évaluation
- Deuxième Semaine Du Développement Embryonnair1Document9 pagesDeuxième Semaine Du Développement Embryonnair1mioioPas encore d'évaluation
- Première: DéveloppementDocument25 pagesPremière: DéveloppementDjaa MilaaPas encore d'évaluation
- Controle Neuro-Endocrinien de L - App Gen. Fem. (C.M.)Document21 pagesControle Neuro-Endocrinien de L - App Gen. Fem. (C.M.)DrissPas encore d'évaluation
- Appareil Reproducteur FemininDocument14 pagesAppareil Reproducteur FemininDrissPas encore d'évaluation
- Reproducteur Féminin (C.M.)Document26 pagesReproducteur Féminin (C.M.)DrissPas encore d'évaluation
- CoursDocument16 pagesCoursKoki kokiPas encore d'évaluation
- Cours 1 - Structure de L - ADN PDFDocument47 pagesCours 1 - Structure de L - ADN PDFFowzia PatelPas encore d'évaluation
- QCM Microbio T2 +corrigéDocument20 pagesQCM Microbio T2 +corrigéIkram La100% (3)
- Définitions Des GlucidesDocument6 pagesDéfinitions Des Glucidesأحمد حمانPas encore d'évaluation
- 07-Le Tissu NerveuxDocument11 pages07-Le Tissu NerveuxBemmoussat AsmaPas encore d'évaluation
- Banque GénétiqueDocument19 pagesBanque GénétiqueMeryem AnjjarPas encore d'évaluation
- Histoire de La GénétiqueDocument4 pagesHistoire de La GénétiqueSaoussen BenmohamedPas encore d'évaluation
- Les TissuesDocument3 pagesLes TissuessofianePas encore d'évaluation
- Histologie GeneraleDocument203 pagesHistologie GeneraleAnou Kalenga100% (1)
- La Cellule ProcaryoteDocument3 pagesLa Cellule Procaryotea.KPas encore d'évaluation
- 4-La BiocompatibilitéDocument7 pages4-La BiocompatibilitéRania Feriel BlhPas encore d'évaluation
- Introduction Général Biocapteurs-1Document31 pagesIntroduction Général Biocapteurs-1Khadidja100% (1)
- Canevas Socle Commun 1er A SNVDocument37 pagesCanevas Socle Commun 1er A SNVSamedDerdourPas encore d'évaluation
- Tissu SanguinDocument15 pagesTissu SanguinAlaa Sobhi100% (1)
- Emille Pinel Vie Et MortDocument95 pagesEmille Pinel Vie Et Mortzavpix100% (1)
- Division CellulaireDocument2 pagesDivision CellulaireMaya MayaPas encore d'évaluation
- BOUREGAB - 1 - 1696255285155 - 2-La Cellule - Cours - INCDocument3 pagesBOUREGAB - 1 - 1696255285155 - 2-La Cellule - Cours - INCray.abbadiPas encore d'évaluation
- TD Adressage L1SV 2021Document5 pagesTD Adressage L1SV 2021kirito 83Pas encore d'évaluation
- Lefort René - Codex Morbus PDFDocument258 pagesLefort René - Codex Morbus PDFeuqehtbPas encore d'évaluation
- Devoir N°2 SMDocument3 pagesDevoir N°2 SMILIASS GERMANYPas encore d'évaluation
- Introduction A La Bio CellDocument15 pagesIntroduction A La Bio CellLilou GrambertPas encore d'évaluation
- Malek Thèse Final 2013Document195 pagesMalek Thèse Final 2013Certaf MaghnjaPas encore d'évaluation
- Regulation Du Métabolisme PDFDocument2 pagesRegulation Du Métabolisme PDFrainer siegfried simo100% (1)
- Structure Des Bactéries 2018 2019Document8 pagesStructure Des Bactéries 2018 2019Ismail HamrouniPas encore d'évaluation
- Cours BIO-1-Définition & Origine de La VieDocument90 pagesCours BIO-1-Définition & Origine de La VieBenjamin Plee100% (1)
- Chap 2. Interaction Des Biomatériaux Avec L'organismeDocument5 pagesChap 2. Interaction Des Biomatériaux Avec L'organismeAbdou Djelama50% (2)
- UE2 - Biologie Cellulaire - Production D - Énergie Dans La CelluleDocument16 pagesUE2 - Biologie Cellulaire - Production D - Énergie Dans La Cellulesun-nee-chan9Pas encore d'évaluation
- ARN Catalytiques - Édition - VirologieDocument39 pagesARN Catalytiques - Édition - Virologiecoursenligne.lille1100% (2)
- CytobiologieDocument17 pagesCytobiologiedjaouad bourouaisPas encore d'évaluation