Vous aimerez peut-être aussi
- ImmunoDocument29 pagesImmunoJalelo AbdelPas encore d'évaluation
- TD2BMS6 PDFDocument88 pagesTD2BMS6 PDFAmine BeninePas encore d'évaluation
- Lecon - Régulation de L'expression Des Gènes - Pharm2 - Dec8 - 2021Document45 pagesLecon - Régulation de L'expression Des Gènes - Pharm2 - Dec8 - 2021doukarafousseyni7Pas encore d'évaluation
- Cours 7 Régulation de L'expression Des GènesDocument15 pagesCours 7 Régulation de L'expression Des GènesFarah B. BtoushPas encore d'évaluation
- Chapitre 3 La Régulation de L'expression Des GènesDocument16 pagesChapitre 3 La Régulation de L'expression Des GènesalmnaouarPas encore d'évaluation
- Regulation D ADNDocument44 pagesRegulation D ADNYousra NanoPas encore d'évaluation
- 9.2 Régulation de L Expression GénétiqueDocument20 pages9.2 Régulation de L Expression GénétiquedamonsalvatorebadvampsPas encore d'évaluation
- Régulation de L'expression Génique: Acétyltransférase (HAT) : Neutralise La Charge + de La Queue DesDocument3 pagesRégulation de L'expression Génique: Acétyltransférase (HAT) : Neutralise La Charge + de La Queue DesNess LzPas encore d'évaluation
- Ronéo Ronéos Biologie Biologie 4 Fini 2022Document12 pagesRonéo Ronéos Biologie Biologie 4 Fini 2022lialazzaraPas encore d'évaluation
- 3-Régulation de L - Expression Des Gènes N.MDocument3 pages3-Régulation de L - Expression Des Gènes N.Msevda mouviePas encore d'évaluation
- Cours Licence 2 BioDocument18 pagesCours Licence 2 BioGayaPas encore d'évaluation
- REGULATION DE LA TRANSCRIPTION Version 2017Document58 pagesREGULATION DE LA TRANSCRIPTION Version 2017Al-mouzammil Sylla100% (1)
- 6-Transcription Et Maturation Des ARNDocument10 pages6-Transcription Et Maturation Des ARNmed.hmamouch99Pas encore d'évaluation
- 03 - Génétique BactérienneDocument60 pages03 - Génétique Bactériennebouchakour meryemPas encore d'évaluation
- BiochDocument243 pagesBiochyouness.khalfaouiPas encore d'évaluation
- Cours RéglationDocument33 pagesCours RéglationBADzPas encore d'évaluation
- Genetique1an16-Regulation Expression GenesDocument8 pagesGenetique1an16-Regulation Expression GenesAmine BeninePas encore d'évaluation
- Facteurs de TransreptionsDocument6 pagesFacteurs de TransreptionsYahiaab RahmaniPas encore d'évaluation
- Cours 8 - La Synthèse ProtéiqueDocument8 pagesCours 8 - La Synthèse ProtéiqueFarouk Ben YahiaPas encore d'évaluation
- Du Gène À La Protéine 2019-20Document20 pagesDu Gène À La Protéine 2019-20Abdelhamid ArouiPas encore d'évaluation
- BCM 7 Transcription TraductionDocument56 pagesBCM 7 Transcription TraductionJoel AmaniPas encore d'évaluation
- Transcription Et Maturation Des ARNDocument26 pagesTranscription Et Maturation Des ARNMariama Ali houmadiPas encore d'évaluation
- Exposé Du Groupe 9Document35 pagesExposé Du Groupe 9Grâce KABOREPas encore d'évaluation
- Correction Devoir de Génétique MoléculaireDocument3 pagesCorrection Devoir de Génétique MoléculairessggnfPas encore d'évaluation
- Biochimie - Pré-Rentrée - Cours 4 - Du Gène À La Protéine - Errata Dans Le PolyDocument45 pagesBiochimie - Pré-Rentrée - Cours 4 - Du Gène À La Protéine - Errata Dans Le PolyNathan CohenPas encore d'évaluation
- 19 Transcriptional Regulation in Eukaryotes.en.FrDocument8 pages19 Transcriptional Regulation in Eukaryotes.en.FrAbdichakour Med HassanPas encore d'évaluation
- TRAnscriptionDocument16 pagesTRAnscriptionmaysounPas encore d'évaluation
- Chapitre II Cours 1Document20 pagesChapitre II Cours 1Abderahmane KmrPas encore d'évaluation
- Biologie Moléculaire Et GG 2Document30 pagesBiologie Moléculaire Et GG 2Sangaré soungaloPas encore d'évaluation
- BMGG La TranscriptionDocument22 pagesBMGG La TranscriptionAbbou BelkaisPas encore d'évaluation
- Chap 1A3 Expres IG V2Document5 pagesChap 1A3 Expres IG V2NokqtoPas encore d'évaluation
- 3éme Cours Génomique Structurale Et ProtéomiqueDocument17 pages3éme Cours Génomique Structurale Et ProtéomiqueHALIMAPas encore d'évaluation
- 131 Le Systeme Membranaire Interne Divers AuteursDocument47 pages131 Le Systeme Membranaire Interne Divers AuteursmbusaPas encore d'évaluation
- Chapitre 1 G Nie G N Tique Introduction 2 PDFDocument51 pagesChapitre 1 G Nie G N Tique Introduction 2 PDFfatiPas encore d'évaluation
- 1-Diapo 1. ORGANISATION DU GÉNOME - PCEM2Document109 pages1-Diapo 1. ORGANISATION DU GÉNOME - PCEM2oumaimahenteti5Pas encore d'évaluation
- Transcription S4 2012Document45 pagesTranscription S4 2012Flarine RamePas encore d'évaluation
- Med Genetique-Transcription EucaryotesDocument20 pagesMed Genetique-Transcription EucaryotesRacha MalakPas encore d'évaluation
- trace écrite-chapitre 2Document6 pagestrace écrite-chapitre 2yezrouraliPas encore d'évaluation
- Recepteurs Protéines - Partie 3.2Document49 pagesRecepteurs Protéines - Partie 3.2Anonymous MKSfyYyODPPas encore d'évaluation
- UE1 - Biochimie - Expression Du GénomeDocument45 pagesUE1 - Biochimie - Expression Du Génomesun-nee-chan9Pas encore d'évaluation
- III. Mécanismes Moléculaires de L'expression de L'information GénétiqueDocument54 pagesIII. Mécanismes Moléculaires de L'expression de L'information GénétiqueNaima BouhmaidaPas encore d'évaluation
- Transcription 21 - 11 - 2023Document55 pagesTranscription 21 - 11 - 2023Elmahdi BOUHFIDPas encore d'évaluation
- ENDOCRINOLOGIE-2024Document95 pagesENDOCRINOLOGIE-2024mohamedazizhmida000Pas encore d'évaluation
- 16-Traduction de L'information GénétiqueDocument33 pages16-Traduction de L'information GénétiqueInsafPas encore d'évaluation
- Biologie Moléculaire Des Plantes - 8 NovembreDocument16 pagesBiologie Moléculaire Des Plantes - 8 Novembrezuzusmail12Pas encore d'évaluation
- Facteurs de Transcription 2016 BT1Document66 pagesFacteurs de Transcription 2016 BT1Damien VicairePas encore d'évaluation
- La TranscriptionDocument28 pagesLa TranscriptionMelissa BaloulPas encore d'évaluation
- Chapitre I: L'expression de L'information GénétiqueDocument74 pagesChapitre I: L'expression de L'information Génétiqueahméd_facebPas encore d'évaluation
- Chapitre 2 BM23Document50 pagesChapitre 2 BM23Mbaye AbouPas encore d'évaluation
- Récept Intracellulaires-Cours M1-2023Document16 pagesRécept Intracellulaires-Cours M1-2023Aggoun FadouaPas encore d'évaluation
- Biologie Moléculaire TD 03Document29 pagesBiologie Moléculaire TD 03biotech micro100% (1)
- TD4 Régulation Du MétabolismeDocument24 pagesTD4 Régulation Du MétabolismeRoumaissa MerazigPas encore d'évaluation
- Empreinte Dnase Gene RaporterDocument27 pagesEmpreinte Dnase Gene RaporterMaï Ssa100% (1)
- Biologie Moléculaire Chapitre 1 T Procaryote M1Document8 pagesBiologie Moléculaire Chapitre 1 T Procaryote M1اميرة في عرش ابيPas encore d'évaluation
- Génétique - CHAP3 PDFDocument21 pagesGénétique - CHAP3 PDFMɞɞɳ LįghtPas encore d'évaluation
- Biologie Moléculaire Chapitre 3 REGULATION M1 1Document8 pagesBiologie Moléculaire Chapitre 3 REGULATION M1 1اميرة في عرش ابيPas encore d'évaluation
- La Transcription l3 Microbiologie 2022Document8 pagesLa Transcription l3 Microbiologie 2022rou röuPas encore d'évaluation
- Transcription 2017 PDFDocument59 pagesTranscription 2017 PDFAntoine MoitiéPas encore d'évaluation
- Ladressage Des Protéines - PDF Dans RERDocument48 pagesLadressage Des Protéines - PDF Dans RERamal ELYOUBIPas encore d'évaluation
- VRD 6Document43 pagesVRD 6Nesrine RAPas encore d'évaluation
- Medecine Batna-ProgramDocument65 pagesMedecine Batna-ProgramNesrine RAPas encore d'évaluation
- 2-2-La Gestion Des Salles de SciencesDocument8 pages2-2-La Gestion Des Salles de SciencesNesrine RAPas encore d'évaluation
- Series TD BIM 2015Document29 pagesSeries TD BIM 2015Nesrine RA100% (1)
- Cours BIB2014Document41 pagesCours BIB2014Nesrine RAPas encore d'évaluation
- M.T 041 1Document89 pagesM.T 041 1Taim KhouriPas encore d'évaluation
- Gaz NaturelDocument8 pagesGaz NaturelLyn DA100% (1)
- Rahou HananDocument55 pagesRahou HananALL about ALLPas encore d'évaluation
- Devoir de Contrôle N°2 - SVT - 3ème SC Exp (2009-2010)Document3 pagesDevoir de Contrôle N°2 - SVT - 3ème SC Exp (2009-2010)sonilola100% (1)
- La Nutrition Azotée Chez Les Végétaux SupérieursDocument10 pagesLa Nutrition Azotée Chez Les Végétaux Supérieursحبيبي اللهPas encore d'évaluation
- Partie2 DEFDocument49 pagesPartie2 DEFDjamel HamoudiPas encore d'évaluation
- L'enfant Au Tambour V2Document4 pagesL'enfant Au Tambour V2JurjePas encore d'évaluation
- QCM Diriges Proteines Acides AminesDocument5 pagesQCM Diriges Proteines Acides AminesSaboPas encore d'évaluation
- TP2 BCH 221 Cholestérol 2Document4 pagesTP2 BCH 221 Cholestérol 2joellemarie001Pas encore d'évaluation
- Cuadernillo Estudio Resumido Fundamentos Organica Jun2020Document150 pagesCuadernillo Estudio Resumido Fundamentos Organica Jun2020Ivette SánchezPas encore d'évaluation
- NJAMPOU PhilippeDocument66 pagesNJAMPOU PhilippeCedric GamegnePas encore d'évaluation
- Etude 2 - Coussinet Dans Cage MétalliqueDocument7 pagesEtude 2 - Coussinet Dans Cage MétalliqueYoussef HidanePas encore d'évaluation
- Tahri Ahmed - 3Document136 pagesTahri Ahmed - 3Daouda DIAGNEPas encore d'évaluation
- Spé SVTDocument8 pagesSpé SVTanissaPas encore d'évaluation
- LEXIQUE MULTILINGUE de La Vigne Et Du Vin Éric GlatreDocument400 pagesLEXIQUE MULTILINGUE de La Vigne Et Du Vin Éric GlatreMarta VarekaiPas encore d'évaluation
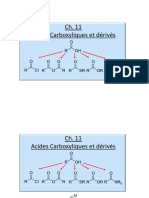
- Ch11 Acides Esters AmidesDocument56 pagesCh11 Acides Esters AmidesYugi kevin14Pas encore d'évaluation
- Liste Des Pesticides Homologues 2016Document48 pagesListe Des Pesticides Homologues 2016habib ibn dkhilPas encore d'évaluation
- 3A BT MCS - Matières PlastiquesDocument6 pages3A BT MCS - Matières PlastiquesFélix KouassiPas encore d'évaluation
- 11 AlcènesDocument49 pages11 AlcènesgaPas encore d'évaluation
- TP4 - Mo PDFDocument3 pagesTP4 - Mo PDFAimen Kechout0% (1)
- Chimie Des Polymeres Exercices Et Problemes Corriges 2 Ed - SommaireDocument22 pagesChimie Des Polymeres Exercices Et Problemes Corriges 2 Ed - Sommairezina allaouaPas encore d'évaluation
- ChapIII Bioch 2020Document60 pagesChapIII Bioch 2020dohaPas encore d'évaluation
- TD Substitution Nucleophile Corrige 4Document6 pagesTD Substitution Nucleophile Corrige 4Daniel Nd100% (1)
- VD Lemoine HuguesDocument250 pagesVD Lemoine HuguesibrahimaPas encore d'évaluation
- Insecticides-Fongicides Des Méthodes Naturelles Pour Protéger Les Plantes Contre Les RavageursDocument6 pagesInsecticides-Fongicides Des Méthodes Naturelles Pour Protéger Les Plantes Contre Les Ravageurskenfack sergePas encore d'évaluation
- LoldedrDocument72 pagesLoldedrLilium CattleyaPas encore d'évaluation
- TD de Milieux de DepotsDocument5 pagesTD de Milieux de DepotsTAKOUGOUM CharlePas encore d'évaluation
- MémoireDocument58 pagesMémoireIkram MellahPas encore d'évaluation
- Proprietes Saccharose PDFDocument34 pagesProprietes Saccharose PDFslimaniPas encore d'évaluation
- Rappels Indispensables de GénétiqueDocument10 pagesRappels Indispensables de Génétiquebakiclio5Pas encore d'évaluation