Académique Documents
Professionnel Documents
Culture Documents
Reviser Son Bac Avec Le Monde SCIENCES de LA VIE
Transféré par
itcomp_2011Copyright
Formats disponibles
Partager ce document
Partager ou intégrer le document
Avez-vous trouvé ce document utile ?
Ce contenu est-il inapproprié ?
Signaler ce documentDroits d'auteur :
Formats disponibles
Reviser Son Bac Avec Le Monde SCIENCES de LA VIE
Transféré par
itcomp_2011Droits d'auteur :
Formats disponibles
T
erm
S
LESSENTIEL DU COURS
Des ches synthtiques
Les points et dnitions cls
Les repres importants
DES SUJETS DE BAC
Des questions types
Lanalyse des sujets
Les raisonnements
Les plans dtaills
Les piges viter
DES ARTICLES DU MONDE
Des articles du Monde
en texte intgral
Un accompagnement
pdagogique de chaque
article
UN GUIDE PRATIQUE
La mthodologie
des preuves
Astuces et conseils
H
O
R
S
-
S
R
I
E
H
o
r
s
-
s
r
i
e
L
e
M
o
n
d
e
,
a
v
r
i
l
2
0
1
3
Rviser son bac
avec
N
o
u
v
e
a
u
x
p
r
o
g
r
a
m
m
e
s
2
0
1
3
Sciences de la vie et
de la Terre
:
H
I
K
P
O
A
=
\
U
\
^
U
U
:
?
k
@
a
@
a
@
c
@
p
"
M
0
5
4
0
7
-
2
H
-
F
:
7
,
9
0
E
-
R
D
En partenariat avec
Avec la collaboration de :
Manon Corbin
Marie-Nol Morin-Ganet
Sandrine Henry
Sciences de la vie et de la Terre T
erm
S
(nouveaux programmes)
Une ralisation de
En partenariat avec
Rviser son bac
avec
r
u
e
d
e
s
c
o
l
e
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
i
o
n
,
d
i
f
f
u
s
i
o
n
e
t
c
o
m
m
u
n
i
c
a
t
i
o
n
s
t
r
i
c
t
e
m
e
n
t
i
n
t
e
r
d
i
t
e
s
r
u
e
d
e
s
c
o
l
e
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
i
o
n
,
d
i
f
f
u
s
i
o
n
e
t
c
o
m
m
u
n
i
c
a
t
i
o
n
s
t
r
i
c
t
e
m
e
n
t
i
n
t
e
r
d
i
t
e
s
LESSENTIEL DU COURS
GNTIQUE ET VOLUTION p. 5
chapitre 01 Le brassage gntique et sa contribution
la diversit gntique p. 6
chapitre 02 Diversication gntique et diversication
des tres vivants p. 12
chapitre 03 De la diversication des tres vivants
lvolution de la biodiversit p. 18
chapitre 04 Un regard sur lvolution de lhomme p. 24
chapitre 05 Les relations entre organisation et mode
de vie, rsultat de lvolution : lexemple de la vie xe
chez les plantes p. 30
LE DOMAINE CONTINENTAL ET SA DYNAMIQUE p. 37
chapitre 06 La caractrisation du domaine continental :
lithosphre continentale, reliefs et paisseur crustale p. 38
chapitre 07 Contexte de la formation des chanes
de montagnes et disparition des reliefs p. 44
chapitre 08 Le magmatisme en zone de subduction : une
production de nouveaux matriaux continentaux p. 50
ENJEUX PLANTAIRES CONTEMPORAINS p. 55
chapitre 09 Gothermie et proprits thermiques de la Terre p. 56
chapitre 10 La plante domestique p. 60
LE MAINTIEN DE LINTGRIT DE LORGANISME :
QUELQUES ASPECTS DE LA RACTION IMMUNITAIRE p. 65
chapitre 11 La raction inammatoire, un exemple
de rponse inne p. 66
chapitre 12 Limmunit adaptative, prolongement
de limmunit inne p. 72
chapitre 13 Le phnotype immunitaire au cours de la vie p. 78
NEURONE ET FIBRE MUSCULAIRE : LA COMMUNICATION NERVEUSE p. 83
chapitre 14 Le rexe myotatique, un exemple de
commande rexe du muscle p. 84
chapitre 15 Motricit volontaire et plasticit crbrale p. 88
LE GUIDE PRATIQUE p. 93
SOMMAIRE
En partenariat avec
Compltez vos rvisions du bac sur www.assistancescolaire.com :
mthodologie, ches, exercices, sujets d'annales corrigs... des outils gratuits et efcaces
pour prparer l'examen.
AVANT-PROPOS
Cet ouvrage, consacr au programme spcique de Sciences de la vie et de la Terre de terminale S et
conu par des professeurs enseignant ce niveau, constitue une prparation originale et efcace lpreuve
crite du baccalaurat en SVT.
Pour vous prparer au mieux au baccalaurat, vous trouverez la n de louvrage, dans le Guide
pratique, la prsentation dtaille de lpreuve de SVT, les exigences des correcteurs et des recommanda-
tions y compris des conseils de mthode et dorganisation pour russir cette preuve.
Dans la double page de Lessentiel du cours, les principales notions de chaque chapitre, accom-
pagnes de schmas, sont expliques et prsentes de manire structure et synthtique, dans le
respect du contenu et de lesprit du programme ofciel de SVT de terminale S. Dans les colonnes,
sont prcisment dnis les mots et les notions cls du cours et des zoom sur certains points du
programme vous permettent dapprofondir vos connaissances.
Dans Unsujet paspas, unsujet complet vous est propos, accompagn de soncorrig et des conseils de
lenseignant sur les principaux piges viter. Pour chaque chapitre, cest lun des trois exercices de la nou-
velle preuve crite de SVTqui est trait. Lensemble des sujets couvre les diffrents exercices de lpreuve
crite de SVT au baccalaurat : partie 1 (restituer ses connaissances lors dune question de synthse ou
dun QCM), partie 2 exercice 1 (raisonner dans le cadre dun problme scientique) et partie 2 exercice 2
(pratiquer une dmarche scientique). Cette rubrique se rvle donc tre un outil efcace pour sentraner
tout au long de lanne lpreuve du baccalaurat.
Pour chaque chapitre, des articles issus du quotidien Le Monde, ont t slectionns pour leur in-
trt et leur pertinence au regard dune ou de plusieurs notions abordes dans le cours. Il sagit
darticles rcents, qui montrent combien les problmatiques des Sciences de la vie et de la Terre
sinscrivent dans lactualit, suscitent des polmiques ou des controverses, que ce soit dans le
domaine de la sant ou dans celui de lenvironnement. Ces articles montrent galement la rapidit de lvo-
lution des connaissances en biologie et en gologie notamment. Les articles choisis vous permettent dap-
profondir les notions ducours, dtayer vos propos lors des preuves, grcedes exemples issus duMonde,
dalimenter votre rexioncritique et, nalement, dapprhender les sciences comme unsavoir toujours en
construction.
Cet ouvrage est une aide prcieuse pour russir lpreuve de SVT, et au-del, pour dcouvrir la
richesse des Sciences de la vie et de laTerre.
M.-N. M.-G.
Edit par la Socit Editrice du Monde 80, boulevard Auguste Blanqui 75013 Paris
Tl : +(33) 01 57 28 20 00 Fax : +(33) 01 57 28 21 21
Internet : www.lemonde.fr
Prsident du Directoire, Directeur de la Publication : Louis Dreyfus
Directeur de la rdaction : Alain Frachon
Imprim par Maury
Commission paritaire des journaux et publications : n 0712C81975
Dpt lgal : mars 2013
Achev dimprimer : mars 2013
Numro hors-srie ralis par Le Monde Le Monde rue des coles 2013.
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
partie
maternelle
cordon
ombilical
vaisseaux
sanguins
ftaux
P
L
A
C
E
N
T
A
sac amniotique
cordon
ombilical utrus
maternel
GNTIQUE
ET VOLUTION
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
6
LESSENTIEL DU COURS
Gntique et volution 7
LESSENTIEL DU COURS
Gntique et volution
Le brassage gntique
et sa contribution la
diversit gntique
Une multitude de gamtes
diffrents
Chezles organismes diplodes, lareproductionsexue
implique la formation de cellules reproductrices,
ou gamtes haplodes, contenant chacune la moi-
ti de linformation gntique de lindividu qui
les produit. lexception des gamtes, chacune
des cellules dun organisme diplode possde des
paires de chromosomes homologues : lun dori-
gine paternelle et lautre dorigine maternelle. La
premire tape de la formation des gamtes est une
division cellulaire appele miose, qui permet dob-
tenir quatre cellules haplodes partir dune cellule
diplode. Elle est prcde d'une copie lidentique
dumatriel gntique (rplicationdelADN). Chaque
chromosome passe alors dune chromatide deux
chromatides surs.Lamioseest unedoubledivision
cellulaire. Lapremire divisionde miose se droule
en plusieurs tapes. Au cours de la prophase I, les
chromosomes se condensent, puis leschromosomes
homologues se placent cte cte : ils sapparient.
En mtaphase I (3), les chromosomes se disposent
sur le plan quatorial, au centre de la cellule. En
anaphase I, les deux chromosomes de chaque
paire se sparent et migrent chacun alatoirement
vers lun des ples de la cellule. Les chromosomes
dorigine paternelle ou maternelle se rpartissent
au hasard, les paires tant indpendantes les unes
des autres. Cest le brassage interchromosomique.
En n de tlophase I, deux cellules haplodes, ne
T
ous les individus appartenant une mme espce pos-
sdent le mme nombre de chromosomes et les mmes
gnes, la mme position le long de l'ADN. Au cours de
lvolution, des mutations peuvent conduire lapparition de
diffrentes versions des gnes : les allles. Diffrents brassages
au cours de la reproduction sexue expliquent que chaque
individu (hormis les jumeaux vrais) soit gntiquement unique,
cest--dire une combinaison unique dallles.
Quantit dADN par cellule
4Q
2Q
Q
Temps
Rplication
de lADN
Sparation des chromosomes
homologues
Sparation des
chromatides surs
Variation de la quantit de matriel chromosomique lors
de la miose
A1
1
B1
A2
B2
X
Y
C1
D1
C2
D2
4
5
C1
D1
A2
B2
C1
D1
A2
B2
A1
B1
C2
D2
A1
B1
C2
D2
Chromatide
Cellule 3 paires
de chromosomes
Formation dune chromatide
sur pour chaque chromosome
par rplication de lADN
Rpartition des chromosomes
au centre de la cellule.
Les chromosomes de chaque paire
se rpartissent alatoirement dun ct
ou de lautre du plan quatorial Chromosome
Chr omatide
sur
Gne A
Gne B
Allles du gne A
Allles du gne B
2
Sparation de la cellule en deux cellules
3 chromosomes 2 chromatides
(un de chaque paire)
Sparation des chromatides surs
dans chaque cellule issue de la premire
division de miose
La deuxime division de miose aboutit
4 cellules possdant chacune un chromosome
de chaque paire une chromatide
3
portant plus quun seul chromosome de chaque
paire, sont formes (4).
La seconde division de miose ressemble une
mitose : les deux chromatides surs de chaque
chromosome sont spares. Une cellule en miose
donnera donc quatre cellules diffrentes, conte-
nant une copie de chaque gne, soit lallle dori-
gine paternelle, soit lallle dorigine maternelle.
Les chromosomes homologues sont si troitement
accols lors de la prophase I que des changes de
matriel gntique peuvent se produire entre chro-
matides : cest le brassage intrachromosomique.
Au hasard, les chromatides chevauchantes se cou-
pent, puis le bras coup se ressoude avec lautre
chromatide. Cest le crossing-over.
Ainsi, partir dune cellule mre de gamtes mles,
on obtient quatre spermatozodes gntiquement
diffrents. Pour les gamtes femelles, les divisions
sont ingales car seule une cellule garde tout le
cytoplasme ; les autres dgnrent. Si dans les
gamtes mles la miose est acheve, pour les
gamtes femelles, la miose est arrte en cours
de processus et sachvera aprs la fcondation
(avant la fcondation on parle donc dovocyte et
non dovule).
Le hasard de la fcondation
Les brassages au cours de la miose permettent
dobtenir une grande diversit de gamtes conte-
nant chacun une combinaison unique et nouvelle
dallles. Chez chaque parent, un seul de ces ga-
mtes sera impliqu dans la fcondation.
Dans une fratrie humaine, la probabilit que deux
individus nonjumeauxvrais soient gntiquement
identiques est quasiment nulle.
Dans lespce humaine, la migration indpendante
des chromosomes (brassage interchromosomique)
permet thoriquement la formation de 2
23
types
de gamtes diffrents et la rencontre au hasard
avec un gamte de lautre sexe conduit chaque
fcondation (2
23
)
2
, soit 70 000 milliards de combi-
naisons possibles pour une cellule-uf ou zygote.
Ce chiffre est trs sous-valu car il ne tient pas
compte du brassage intrachromosomique d aux
crossing-over. Une quasi-innit de combinaisons
gntiques diffrentes sont en ralit possibles.
Toutefois, seule une fraction de ces zygotes est
viable et peut poursuivre son dveloppement. Une
anomalie chromosomique peut par exemple em-
pcher le droulement des divisions de lembryon
et conduire un avortement spontan.
Les anomalies au cours de la
miose : sources de troubles
mais aussi de diversication
Lors de la miose, deux chromosomes (miose I)
ou deux chromatides (miose II) peuvent migrer
vers le mme ple de la cellule. Dans ce cas,
certains gamtes porteront un chromosome de
moins, et dautres un chromosome surnum-
raire. Ces gamtes peuvent tre fcondants, mais
le zygote ne possdera pas le nombre de chromo-
somes spcifique de son espce : sil est viable,
lindividu sera porteur dune anomalie chromoso-
mique, comme une monosomie (45 chromosomes
au lieu de 46) ou une trisomie (47 chromosomes).
Lors des crossing-over, des morceaux de chroma-
tides peuvent tre changs de faon ingale : les
gamtes qui rsultent de ces divisions peuvent
soit ne plus porter certains gnes (on parle de
dltion de gnes), soit en avoir deux copies (on
parle alors de duplication de gnes). Si la fcon-
dation implique un gamte portant l'une de ces
anomalies, lembryon peut ne pas survivre ou
prsenter des malformations dues cette anoma-
lie gntique. Dans certains cas, la duplication de
gnes a permis de crer des familles multig-
niques, comme les gnes du complexe majeur
dhistocompatibilit (CMH) ou les gnes codant
pour les globines.
Anomalie de rpartition des chromosomes homologues
enpremire divisionde miose : la moiti des cellules lles
aura unchromosome surnumraire, lautre moiti aura un
chromosome en moins
MOTS CLS
ALLLES
Versions diffrentes dun mme
gne, caractrises par de lgres
diffrences au niveau de leur s-
quence nuclotidique.
CHROMOSOMES
HOMOLOGUES
Chromosomes appartenant une
mme paire dans le caryotype
dune espce diplode. Ils portent
les mmes gnes, au mme locus,
mais pas forcment les mmes
allles.
DIPLODE (2N)
Qui possde deux lots de chromo-
someshomologues, cest--diredes
paires de chromosomes.
GAMTE
Cellule reproductrice haplode
(ovule chez la femelle, spermato-
zode chez le mle).
HAPLODE (N)
Qui ne possde quun seul lot de
chromosomes (une seule version
de chaque gne).
ZOOM SUR
LE BRASSAGE
INTERCHROMOSOMIQUE
Il correspond la migration ind-
pendante des diffrentes paires
de chromosomes au cours de la
premire division de miose. On
peut le mettre envidence enana-
lysant les rsultats de croisements
tudiant des gnes ports par des
pairesdechromosomesdiffrentes
(gnes indpendants).
LE BRASSAGE
INTRACHROMOSOMIQUE
Il augmente la diversit des ga-
mtes enformant des chromatides
recombines par crossing-over en
prophase de premire division de
miose. Les gamtes de type re-
combin possdent une nouvelle
combinaison dallles quaucun
des parents ne prsentait. Onpeut
mettre en vidence ce brassage en
analysant les rsultats de croise-
ments tudiant des gnes ports
par une mme paire de chromo-
somes (gnes lis).
ZOOM SUR
LES ANOMALIES
CHROMOSOMIQUES
Des perturbations dudroulement
de la miose peuvent conduire
des anomalies chromosomiques
dont quelques-unes peuvent tre
viables : trisomie 21, trisomie 18
(mortalit avant lge de 1 an) ou
trisomie13(dcs inuteroouavant
3mois). Ellespeuvent aussi toucher
les chromosomes sexuels : triso-
mieXXY(syndromedeKlinefelter,
homme strile), trisomie XYY, tri-
somie XXXoumonosomie X(syn-
drome de Turner, femme strile).
LES FAMILLES
MULTIGNIQUES
Certains ensembles de gnes
se caractrisent par de grandes
similitudes au niveau de leurs
squences. Ces gnes sont situs
des locus diffrents sur un
mme chromosome ou sur des
chromosomes diffrents. Une res-
semblance de plus de 20 % nest
pas le fait du hasard, mais dune
parent. Ces gnes rsulteraient
de duplications partir dun gne
ancestral. Ce mcanisme, associ
des mutations qui se produisent
de manire indpendante sur les
diffrents duplicatas, permettrait
dexpliquer lexistencedes familles
multigniques. Laduplicationdun
gne constitue une source de di-
versication car la protine peut
acqurir denouvellesfonctionspar
mutations du gne dupliqu, sans
que le gne et la protine dorigine
ne soient affects.
Lexemple de la famille des glo-
bines chez lhomme.
Lhmoglobine est une protine
constitue de lassociation de
quatre chanes polypeptidiques.
Au cours de sa vie, lorganisme
humain fabrique plusieurs types
dhmoglobines, constitues de
chanes de globines diffrentes :
chanes zta, epsilon, alpha, bta,
gamma ou delta. Les six gnes co-
dant pour ces globines sont situs
sur des locus diffrents sur les
chromosomes 16 et 11. La com-
paraison des squences dacides
amins des diffrentes globines
permet de constater leur grande
ressemblance.
UN ARTICLE DU MONDE CONSULTER
Laventure de lembryon Les chemins du
commencement p. 10-11
(CatherineVincent, 5 janvier 1999)
Enjambement des chromatides change de matriel gntique
Une recombinaison homologue en prophase de premire division de miose
Le brassage intrachromosomique
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
LESSENTIEL DU COURS
8
UN SUJET PAS PAS
9 9 Gntique et volution Gntique et volution
Le QCM
Cochez la proposition exacte pour chaque question
de 1 6.
1. Cette photographie reprsente une cellule :
a) 2n = 24, en anaphase dune mitose.
b) 2n = 24, en anaphase I dune miose.
c)2n = 12, en mtaphase dune mitose.
d) 2n = 12, en anaphase II dune miose.
2. Lamitose :
a) est source de diversit gntique.
b) donne naissance 4 cellules partir dune cellule.
c) conserve toutes les caractristiques du caryotype.
d) permet la production des gamtes.
3. Lamiose produit :
a) 4 cellules haplodes partir dune cellule diplode.
b) 2 cellules diplodes partir dune cellule diplode.
c) 4 cellules diplodes partir dune cellule diplode.
d) 2 cellules haplodes partir dune cellule diplode.
4. Larplicationde lADNalieu:
a) entre les deux divisions de la miose.
b) uniquement avant une mitose.
c) uniquement avant une miose.
d) avant la premire division de la miose
5. Lors dune miose se droulant sans anomalie, il
peut seffectuer :
a) un brassage intrachromosomique entre chromo-
somes non homologues.
b) un brassage interchromosomique entre chromo-
somes homologues.
c) unbrassageinterchromosomiquepuis unbrassage
intrachromosomique.
d) unbrassageintrachromosomiquepuis unbrassage
interchromosomique.
Le document suivant reprsente le caryotype dun
enfant atteint dune anomalie chromosomique.
6. Le caryotype ci-dessous peut avoir pour origine :
a) une duplication du chromosome 21 lors de la
miose.
b) une non-disjonction de la paire chromosomique
n 21 lors de la division I de la miose.
c) une non-disjonction de la paire chromosomique
n 21 lors de la division II de la miose.
d) un accident gntique uniquement lors de la
formation des gamtes femelles.
Le sujet de question de synthse
La diversit du vivant a pour origine de nombreux
mcanismes dont certains sont dorigine gntique.
Enprenant comme exemple la transmissionde deux
gnes lis, montrez comment lors de lamiose il peut
seffectuer unbrassage de linformationgntique.
Votre expos sera accompagn de schmas.
Le corrig
QCM : 1. b), 2. c), 3. a), 4. d), 5. d) 6. b)
Lanalyse du sujet de question
de synthse
La prcision deux gnes lis permet de limiter
le sujet au brassage intrachromosomique. Il sagit
alors de trouver un gnotype parental pertinent
(htrozygote) pour deux couples dallles ports
par la mme paire de chromosomes, puis de re-
prsenter les phases de la miose appropries, en
insistant sur diffrents moments de la prophase I
(crossing-over).
Proposition de corrig
Introduction : La miose est dnie comme la divi-
sioncellulaire permettant la productionde gamtes
haplodes ne portant quunallle pour chaque gne
sur les deux que possde le parent.
Problmatique gnrale : Comment expliquer que
des parents ne puissent engendrer que des descen-
dants gntiquement diffrents ?
Problmatique limite au sujet : Dans le cas de
gnes lis, comment expliquer que des descendants
Partie 1 : La miose
puissent porter des associations dallles diffrentes
de leurs parents ?
Point de dpart : Une cellule souche de gamte dun
parent htrozygote pour deux gnes ports par la
mme paire de chromosomes.
I. Deuxgnes lis sont censs migrer ensemble lors
de lanaphase I de la miose
Rsultat : on peut former deux types de gamtes
seulement concernant les deux gnes tudis.
II. Les rsultats exprimentauxrvlent des recom-
binaisons
Les analyses de croisements effectus enlaboratoire,
chez la drosophile par exemple, montrent que les
parents htrozygotes produisent en ralit quatre
types de gamtes, mais dans des proportions qui
ne sont pas quivalentes : une majorit de
gamtes parentaux et une minorit de
gamtes recombins qui portent des
associations dallles qui nexistaient pas
chez le parent.
III. Les changes entre chromosomes ho-
mologues
Les recombinaisons ont lieu en prophase I
de miose, au moment de lappariement
des chromosomes homologues. Des cros-
sing-over se forment et les chromosomes
changent des portions de chromatides :
cest le brassage intrachromosomique. Ce-
pendant, pour uncouple de gnes donn, les
recombinaisons homologues nont lieu que
dans un certain pourcentage des mioses
au moment de la formation des gamtes.
Conclusion : Ltude de croisements concer-
nant uncoupledegnes lis permet demettre
envidencelebrassageintrachromosomique,
qui augmente la diversit possible des ga-
mtes. Si lon considre lensemble des chro-
mosomes et des gnes dunindividu, la com-
binaisondubrassageintrachromosomiqueet
dubrassageinterchromosomique(migration
indpendante des diffrentes paires de chro-
mosomes en miose I) permet de produire
une innit de gamtes diffrents. La fcon-
dationruniraauhasarddeuxgamtes parmi
de nombreuses possibilits.
Ce quil ne faut pas faire
Cocher plusieurs rponses par question dans le
QCM: une seule est exacte.
Confondre brassage interchromosomique
(mtaphase et anaphase I) et brassage
intrachromosomique (prophase I).
Ngliger les schmas ou reprsenter
des chromosomes trop petits.
Se tromper en plaant les allles sur les
chromosomes de la cellule parentale de dpart.
Traiter la fois le brassage interchromosomique
et le brassage intrachromosomique, alors que le
sujet ne porte que sur ce dernier (gnes lis).
Traiter la fcondation alors que le sujet
ne porte que sur la miose.
AUTRES SUJETS POSSIBLES AU BAC SUR CE THME
Partie 1 : Synthse sans documents
Discutez des consquences possibles des anomalies survenues lors de la miose et de la fcondation. (schmas
attendus)
Partie 2.1 : Rdig (tableaucomplter)
Ordonnez et annotez des clichs dobservationde diffrentes phases de la miose.
Partie 2.1 : QCMavec document
tude dungraphique reprsentant lvolutionde la quantit dADNaucours de la miose et de la fcondation.
Une cellule danthre
de lys endivision
NOTIONS CLS
CONVENTIONS
DCRITURE
EN GNTIQUE
Engntique, les diffrents allles
dun gne sont dsigns par une
ou deux lettres.
Lallle dominant est crit en ma-
juscules ou porte le signe (+) en
exposant, lallle rcessif est crit
en minuscules ou ne porte pas
de signe. Ex. : chez la drosophile,
les ailes peuvent tre vestigiales
(allle vg) ounormales (allle vg
+
).
Le phnotype scrit entre cro-
chets. Ex. : [vg
+
] signie que la dro-
sophile porte des ailes normales.
Le gnotype scrit entre paren-
thses. Les deux allles que pos-
sde lindividu diplode sont s-
pars par deuxtraits dont chacun
reprsente lundes chromosomes
homologues. Ex. : le gnotype
dune drosophile htrozygote
scrit (vg
+
//vg).
CROISEMENT TEST,
TEST CROSS
Croisement dunindividuprsen-
tant le phnotype dominant avec
un homozygote rcessif, cest--
dire porteur des allles rcessifs
des gnes considrs. Le croise-
ment test permet, par ltude du
phnotype des descendants, de
mettre enlumire le gnotype des
gamtes du parent prsentant le
phnotype dominant.
DOMINANCE/
RCESSIVIT/
CODOMINANCE
Lorsque les deux chromosomes
homologues portent des allles
diffrents pour un mme gne,
celui qui sexprime au niveau
du phnotype est appel allle
dominant . Lautre est rcessif.
Si les deux allles sexpriment,
comme dans lexemple dugroupe
sanguin AB, ils sont dits codo-
minants .
HOMOZYGOTE/
HTROZYGOTE
Un individu peut possder deux
allles identiques pour un gne :
il est alors homozygote pour ce
gne. Sil possde deuxallles dif-
frents, il est htrozygote.
ZOOM SUR...
La drosophile, organisme
modle en gntique
Lamouchedrosophileest devenue
un organisme modle en gn-
tiquedepuis ledbut duXX
e
sicle.
Elle prsente notamment lavan-
tagedenavoir quunpetit nombre
de chromosomes. Les rsultats
obtenus ont pu tre extrapols
aux autres organismes. Ce sont
notamment les travauxsur la dro-
sophile qui ont permis de faire le
lien entre les chromosomes et les
caractres hrditaires.
DATES CLS
Lhistoire de la gntique
1866 : Mendel, prtre et bota-
niste, labore les premires lois
de la transmission des caractres
hrditaires.
1879 : Fleming observe le com-
portement des chromosomes au
cours de la mitose.
1888 : Strasburger observe le
comportement des chromosomes
au cours de la miose.
1901 : DeVries introduit lanotion
de mutation.
1902 : Suttonet Boveri proposent
que les chromosomes soient les
porteurs de linformation gn-
tique.
1906 : Johanssen introduit le
terme gne pour dsigner un
facteur hrditaire.
1915: Morganapporteles preuves
exprimentales de lathorie chro-
mosomique de lhrdit grce
ses tudes sur les drosophiles.
1941 : Beadleet Tatumtablissent
quun gne est responsable de la
synthseduneenzyme(protine).
1944 : Avery, McLeod et McCarty
dcouvrent que lADN est le sup-
port de linformation gntique
chez la bactrie.
1953: Watsonet Cricklaborent le
modle de la structure en double
hlice de lADN grce aux travaux
de Franklin.
1959 : Lejeune met en vidence
lanomalie chromosomique de la
trisomie 21 (syndrome de Down).
1988 : lancement du projet G-
nome humain.
2003 : achvement du squen-
age complet dugnome humain.
LESSENTIEL DU COURS UN SUJET PAS PAS
A1 A1
B1 B1
A2
B2
A2
B2
A1
B1
A1
B1
A1 A1 A2 A2
B1 B1 B2 B2
A2 A2
B2 B2
2 couples
dallles
2 gnes lis
Gne A :
allles A1 et A2
Gne B :
allles B1 et B2
Cellule souche de gamte
du parent htrozygote
I. Deux gnes lis migrent ensemble
lors de lanaphase I de la miose.
Tlophase I
Tlophase II
A2
B2
A1
B1
A1
B2
A2
B1
A1 A2 A1 A2
B1 B1 B2 B2
Prophase I
crossing-over
Fin de
Prophase I
A1 A2 A1 A2
Les changes entre
chromosomes homologues
Chromatides parentales
gamtes parentaux
Chromatides recombines
gamtes recombins
B1 B1 B2 B2
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
10
L'ARTICLE DU
11 11
Laventure de lembryon
Les chemins du commencement
partir dune seule cellule fconde, un tre humain se fabrique en neuf mois, grce
un processus dune complexit inoue. Dans une enqute en cinq volets, Le Monde
raconte lhistoire extraordinaire de la vie avant la naissance.
C
roissez et multi-
pliez , dit le message
biblique. Et sil sagis-
sait en vrit dun message bio-
logique ? Depuis que la vie est
apparue, observe Jean-Paul Lvy,
elle refait de la vie. Les humains
font dautres humains, comme
tout ce qui vit depuis plus de trois
milliards dannes sest occup
refaire du semblable. Mde-
cin, directeur de lInstitut Cochin
de gntique molculaire, il ne
connat sil en faut un qu un
seul miracle : lmergence, au
sein de la soupe chimique ori-
ginelle, de la premire cellule.
Paroledhommedescience, parole
de raison. Et pourtant ! Comment
ne pas smerveiller, et smer-
veiller encore, devant le gnie du
vivant ? Un processus dune com-
plexit inoue, qui, partir dune
seule cellule fconde, fabrique en
neuf mois un tre humain fait de
chair et de sang, de cur et de sa-
gesse, de sensations et de pense ?
Untre humainpareil auxautres,
et toujours unique ? Car cest bien
l, si lon ose rpondre au savant,
le second miracle de la re-
production. Refaire du semblable,
certes, mais en imposant lappa-
rition du diffrent, seul garant de
ladaptation au milieu. Lespce
doit se maintenir, mais elle doit
aussi tre prte voluer quand
lenvironnement change. Si les
premiers prhominiens, quelques
millions dannes avant nous, ne
staient pas adapts la savane
parce que leur squelette ntait
plus le mme que celui de leurs
cousins forestiers, il nyaurait pas
dhommes aujourdhui , avance
Jean-Paul Lvy. Se conserver et se
transformer en mme temps : ni
la scissiparit (reproduction par
simple divisionde lorganisme), ni
la parthnogense (reproduction
sans fcondation), ni le bouturage
nesavent lefaire. Ils nedonnent
quelques mutations prs quedes
copies de loriginal. Cest pourquoi
la nature a invent la sexualit. Et,
avec elle, la diversit.
Dupremier uf, issucommenous
tous de lafusiondune entit mle
et dune entit femelle, que sait-
on ? Quil se forma voil deux
trois milliards dannes, et quil
bouleversa lordre de la vie. On
peut samuser imaginer linsigni-
anceapparentedecettepremire
rencontre : laccolement de deux
cellules ou de deux molcules
quelconques parmi des milliards
dautres. Un vnement micros-
copique, quasi invisible, et sans
doute la plus grande rvolution
que la Terre ait jamais connue ,
commenteAndrLanganey, biolo-
giste des populations auMusum
national dhistoire naturelle.
lquation un donne deux de
la multiplicationse substitue celle
de la procration. Non pas deux
donnent un , mais plutt un
plus un donnent un autre . Sil
faut semettredeuxpour faireun
troisime, cest, justement, pour
que ce dernier ne soit identique
aucun des premiers. L est la
justification de la poursuite
ternelle de lautre, cette srie de
cycles olespce se divise chaque
fois en lments opposs, chaque
fois destins sunir nouveau ,
ajoute Franois Jacob, prix Nobel
de mdecine et professeur auCol-
lge de France. Linnovation est
dautant plus blouissante quelle
met enjeudes mcanismes dune
prodigieuse ingniosit.
Parce que tout se joue enquelques
instants, parce qu lunit de
temps sajoute celle du lieu, parce
que sy mle, enn, lamour char-
nel dedeuxtres, nousdatonslori-
gine de notre identit aumoment
de la fcondation. Cest oublier la
divisionappelemiose. Certes,
la fusionqui se produit alors entre
spermatozode et ovule, entre le
don du pre et celui de la mre,
lguera lenfant une subtile mo-
saque de leurs gnes respectifs.
Mais la source de la diffrence
se situe bien en amont de cet
vnement. Elle prend naissance
une gnration plus tt, dans les
embryons mmes des futurs pa-
rents. Les cellules sexuelles mles
et femelles les gamtes tmoi-
gnent en effet de leur existence
ds les premires semaines de la
vie in utero. Niches dans les go-
nades, elles yattendent leur heure
pour subir une division nulle
autre pareille la miose , sans
laquelle la diversit des espces
ne serait rien. Comme toutes les
cellules du corps, ces cellules ger-
minales naissent diplodes : elles
contiennent 23 paires distinctes
de chromosomes, chacune pro-
venant pour moiti du pre et
pour moiti de la mre. La miose
a pour rle de rduire de moiti
le nombre de chromosomes des
cellules sexuelles, de les rendre
haplodes. Ainsi pourront-elles,
lorsquelles fusionneront dans la
fcondation, apporter chacune
leur lot gntique, sans que luf
excde pour autant les 23 paires
chromosomiques ncessaires
cest ainsi sondveloppement.
Ignorant le processus de repro-
duction habituel (la mitose, au
cours de laquelle la cellule mre
double sonpatrimoine gntique,
puis se scinde en deux, restituant
un jeu complet de chromosomes
chacune de ses deux cellules
lles), la miose impose ainsi au
spermatozode et lovule deux
divisions successives. Mais elle
fait plus encore. Chorgraphe
dun dlicat ballet biologique, elle
impose aux gnes des gures in-
dites. Empchant par lmmeque
chaque cellule sexuelle nemporte
avec elle, en bloc, une moiti des
gnes dun parent que lui-mme
aurait reu en bloc de lun des
siens. Sil en tait ainsi, chaque
individu hriterait dune moiti
dinformation venant dun des
grands-parents maternels et
lautre dun des grands-parents
paternels, et plus rien des deux
autres , remarque Jean-Paul Lvy.
Heureusement, les chromosomes
ne sont pas indestructibles. Ils
peuvent se casser, se recoller,
bref : se rinventer. Cest ce qui se
produit lors de lamiose. Les deux
chromosomes dune mme paire
commencent par saccoler gne
gne, sur toute leur longueur,
puis, scartant partir du centre,
amorcent la sparation. Mais cer-
taines zones de contact rsistent ;
les gnes jumeaux sentremlent,
shybrident, sapproprient un peu
lun de lautre avant de sloigner
dfinitivement. Cest l, durant
cette valse-hsitation, qua lieu
leur recombinaison , grce
laquelle les cellules sexuelles ne
transmettent pas seulement les
gnes reus dupre oude la mre,
mais un patchwork des deux.
Ainsi, luf fcond hritera dun
patrimoine dont les richesses pro-
viennent au hasard de ses quatre
grands-parents et, travers eux,
des innombrables gnrations
antrieures. Ainsi, la sexualit, en
crant de nouvelles associations
de gnes, autorisera toutes les
audaces de lvolution.
Car peine invent, ce jeu des
possibles entrana une diversi-
cationdes espces sans prcdent.
Comme dans la cosmologie sum-
rienne (o leau, manifestation
primitive de la vie du monde, se
prsente sous un double aspect :
Apsu, eaudouce ouprincipe mle,
et Tiamat, eau sale ou principe
femelle), tous les vers, crustacs,
poissons et plantes qui spanouis-
sent dans la matrice aquatique
originelle sorientent vers leur
contrairepour quesunissent leurs
cellules sexuelles et que se cre
luf qui les perptuera. Comment
ces minuscules gamtes, mis
dans locan par des centaines
despces distinctes, retrouvent-ils
leur moiti ? Les progrs rcents
de limmunologie ont montr
lextrme complication des m-
canismes de reconnaissance du
soi au niveau de lorganisme. Il
est probable que les mcanismes
de reconnaissance du gamte
conjoint ensoient unevariante, ou
peut-tre une version primitive ,
avance Andr Langaney. Avec la
sexualit nat, aussi, la recon-
naissance de lautre. Plus grande
sera la difcult, lorsque les plus
hardis de ces nouveaux habitants
dumonde sessaieront sortir des
eaux. Car les cellules sexuelles ne
peuvent survivre quen milieu
liquide, ce qui explique que les
grenouilles et les salamandres,
contournant le problme, aient
conservleurs amours aquatiques.
Pour crer vritablement le sta-
tut danimal terrestre, la nature,
une fois encore, doit innover. Et
instaurer une mthode simple et
universelle pour que la rencontre
puisse seffectuer hors de leau.
Ce sera la fcondation interne,
lintrieur du corps de la femelle.
Par des moyens variant linni
suivant les espces, le sperme du
mleest ainsi achemin, depuis les
testicules oil seforme, jusquaux
organes o il assure la fconda-
tion. Parfois dans la sduction
anonyme (le mille-pattes sclipse
aprs avoir dpos son sperme
dans une petite toile, dont lodeur
attire irrsistiblement sa belle),
parfois avec autorit, voire ath-
ltisme (chez les collemboles, le
petit mle va jusqu soulever son
norme femelle pour lemmener
au lieu o il a dpos ses sperma-
tophores). Pour transmettre une
gouttelette contenant quelques
millions de spermatozodes, les
variations amoureuses et anato-
miques les plus sophistiques ont
ainsi t essayes. Jusqu ce que,
enn, soit invente la plus efcace
dentre elles : un petit bijou de la
technique baptis pnis, qui a fait
ses preuves depuis des dizaines de
millions dannes chez les reptiles
et les mammifres.
Son principe : un tissu spongieux
capable de se gorger de sang, de
goner et dacqurir la rigidit
ncessaire sapntrationdans le
vagin. lautre extrmit de celui-
ci souvre une deuxime cavit,
lutrus. Il se prolonge de chaque
ct par deux longs conduits, les
trompes de Fallope, dont la partie
terminale (lampoule) saccole
lovaire pour y recueillir lovule
quil met priodiquement.
Cest donc l, au cur du ventre
maternel, quauralieularencontre.
Elle saccomplira au terme dune
course harassante, dispute par
une myriade de spermatozodes
dont unseul sortiragagnant. Celui-
ci, du mme coup, dterminera
le sexe de lenfant natre, selon
que son chromosome sexuel sera
XouY. Garonoulle, le choixest
en effet dict par lappariement
XY ou XX.
La mre ne pouvant produire que
des ovules X, cest donc au pre,
par ses cellules fcondantes X ou
Y, que revient le n mot de laf-
faire. Cest pourquoi les diverses
mthodes exprimentes pour
dterminer et orienter le sexe
des enfants avant leur conception
se fondent sur la sparation des
spermatozodes en groupes X et Y
(latechniquelaplus ablecejour,
rendue publique il y a quelques
mois par une rme amricaine,
consisterendreceschromosomes
uorescents, puis les sparer en
fonctiondeleur luminosit), avant
de fconder lovule par insmina-
tion articielle.
X ou Y : par les voies naturelles,
seul le hasard dcidera. Mais
avant, quel marathon ! Et que
dabandons ! Partis entre 50 et
100 millions, ils ne seront plus
que quelques centaines avoir
franchi lutrus et a avoir remont
jusqu lampoule qui coiffe la
trompe. Le trajet quils ont par-
courir leur prendra prs de deux
heures : 20centimtres, lavitesse
de 50 millionimes de mtre par
seconde. Avec pour tout bagage
un agelle long et mobile, une
belle rserve de carburant et une
petitevsiculebourredenzymes,
lacrosome, che au-dessus de
la tte. Au bout de la compti-
tion : la timbale. Un gigantesque
ovule (0,14 mm de diamtre),
lune des plus grosses cellules
de lorganisme. Lequel nira par
lemporter ? Sur quels mrites,
sur quel exploit sera dsign le
vainqueur ? Nul ne le sait, et nous
sommes tous le produit de cette
loterie. Une certitude, toutefois :
malgr lintense slection subie
par les spermatozodes, lefca-
cit de ce mode de reproduction
est considrable (le record ofciel
pour lespcehumaine, dtenupar
une femme russe du XVIIIe
sicle,
est de soixante-neuf enfants). Et
une piste de recherche : dans cette
rencontre au sommet des voies
utrines, le nez des cellules
semble jouer un rle non ngli-
geable. De mme que les femelles
de multiples espces scrtent des
phromones pour sduire leurs
mles, lovule attire les spermato-
zodes enmettant des substances
chimiques, dont la structure nest
pas sans rappeler celle des mol-
cules odorantes...
Voici donc llu aux portes de son
royaume. Il commence par en
perdre sa coiffe, lacrosome, qui se
dissout et libre ses enzymes. Ces
derniers ont pour missiondatten-
drir la zone pellucide, enveloppe
protectrice qui entoure lovule.
Lequel, ds lultimetapefranchie,
exprime sa satisfaction en met-
tant des milliers depetits granules,
qui empcheront dsormais tout
autre prtendant de le pntrer.
Vient alors le grand moment, la
fusion des deux noyaux mle et
femelle. La naissance de luf,
la restitution de la diplodie. Le
moment zro du dveloppement
embryonnaire.
Dans les vingt-quatre heures qui
suivent sa conception, le futur en-
fant natre se divise en deux cel-
lules. Puis en quatre, puis en huit.
Au stade de trente-deux cellules,
le voil comme une petite mre,
dosonjoli nom, morula . Trs
vite, les cellules sedivisent endeux
groupes, lunpriphrique, lautre
central. La masse cellulaire ex-
terne, appele trophoblaste ,
sera lorigine du placenta ; la
masse interne deviendra lem-
bryon. Lensemble commence
se creuser dune cavit centrale
emplie de liquide.
Empruntant lechemininversedes
spermatozodes, luf redescend
la trompe de Fallope pour gagner
la cavit utrine. Au sixime jour
du dveloppement, cest une
sphre creuse constitue dune
centaine de cellules, le blastocyste,
qui fait son entre dans la cavit
utrine. Un monde dont la paroi,
richement vascularise et place
sous la protection de la progest-
rone, scrte gnreusement fac-
teurs de croissance et lments
nutritifs. Lembryonne tarde pas
sy implanter et senfonce profon-
dment dans lopulent tissu.
Laventure inutero commence.
Catherine Vincent
(5 janvier 1999)
POURQUOI
CET ARTICLE ?
Cet article propose une
bonne synthse et une mise
en perspective des brassages
gntiques au cours de la
miose et de la fcondation.
Il illustre le fait que la re-
production sexue permet
de refaire du semblable
[] en imposant lapparition
du diffrent, seul garant de
ladaptation au milieu : la
plus grande rvolution que
la Terre ait jamais connue.
En effet, ces processus assu-
rent la diversit au sein des
espces mais entranent ga-
lement la diversification du
monde vivant. La deuxime
partie de larticle est hors pro-
gramme, elle renseigne sur
les modes de reconnaissance
des gamtes et lapparition
de la fcondation interne. Le
marathon que constitue
le trajet des spermatozodes
jusqu lovule permet de
rappeler le hasard de la ren-
contre des gamtes, parmi
de nombreuses possibilits.
Gntique et volution Gntique et volution
L'ARTICLE DU
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
12
LESSENTIEL DU COURS
13
LESSENTIEL DU COURS
Gntique et volution Gntique et volution
Diversication gntique
et diversication
des tres vivants
Le transfert horizontal de gnes
Le transfert vertical qualifie la transmission de
matriel gntique dune gnration une autre.
Des transferts horizontaux sont possibles entre des
bactries qui ne descendent pas les unes des autres,
quelles appartiennent ounon la mme espce. Des
gnesdersistanceauxantibiotiquessont ports par
des plasmides, courtes molcules dADN circulaires.
Deux bactries peuvent schanger directement du
matriel gntique plasmidique grce des ponts
cytoplasmiques. Une bactrie acquiert alors la rsis-
tance certains antibiotiques partir dune autre
bactrie. Il sagit dun mcanisme de diversication
rapide. Lusage excessif de traitements antibiotiques
a favoris le dveloppement de populations bact-
riennes de plus en plus rsistantes.
Hybridation et polyplodisation
Deux espces vgtales voisines peuvent tre inter-
fcondes. Leur croisement, naturel ouarticiel, peut
donner des hybrides prsentant des critres varis,
susceptibles dintresser les agriculteurs : produc-
tivit, got, rsistance au froid, un insecte Parmi
les plants issus des graines obtenues partir de ces
croisements seront slectionns ceuxqui expriment
les caractres les plus intressants pour lagriculture.
Les crales destines la consommation, comme
le bl ou le mas, sont issues dune succession de
L
es mutations et la rencontre au hasard de deux gamtes g-
ntiquement uniques ne sufsent pas expliquer la totalit
de la diversication des tres vivants. Celle-ci sest ralise
au cours du temps, depuis les premires formes de vie, il y a
quatre milliards dannes. Les mcanismes mis en jeu sont va-
ris et ne sont pas toujours lis une diversication gntique.
R R
R
R R
R
R
Chromosome bactrien
Copie lidentique du plasmide
portant le gne de rsistance
un antibiotique
Bactrie A rsistante
un antibiotique
Bactrie B sensible
un antibiotique
Pont entre les deux
bactries, permettant
le passage du plasmide
copi
Bactrie A rsistante
un antibiotique
Bactrie B rsistante
au mme antibiotique
que la bactrie A
Plasmide
portant le gne
de rsistance
lantibiotique
Transfert dun plasmide portant un gne de rsistance un antibiotique, dune bactrie rsistante une bactrie non
rsistante (conjugaison)
croisements. Dans le cas de ces vgtaux, il y a eu
augmentation du nombre de chromosomes : on
parledepolyplodisation. Cephnomneest frquent
chez les vgtaux, mais beaucoup plus rare chez les
animaux. Il peut notamment rsulter dune absence
de rduction du nombre de chromosomes lors de la
formation des gamtes.
Lengrain sauvage et lgilope faux peautre sont
diplodes (ils ont 7 paires de chromosomes) et ont
t croiss. Ils ont donn dans leur descendance une
plantettraplode(2 7 paires, soit 28chromosomes),
qui a t elle-mme croise avec un plant diplode.
Cela a donn deux sortes de crales 42 chromo-
somes (3 7 paires). La domesticationa eu plusieurs
effets, telle laugmentation de la taille de lpi mais
galement dunombre de grains par pi. La producti-
vit a donc t amliore.
Linfection par un virus
Prs de 10 % du gnome des mammifres provien-
nent de virus. Ils sont des parasites intracellulaires :
leur gnome sinsre dans celui des cellules infectes.
Cet ajout dematriel gntiquepeut tredfavorable,
mais aussi procurer un avantage transmissible la
descendance sil sagit de cellules germinales.
Des variations dans lexpression
de certains gnes
Charles Darwin au XIX
e
sicle, les Grant au XX
e
sicle
ont tudi les pinsons des Galpagos. Ona dcouvert
plus rcemment que la forme de leur bec tait lie
lexpression de certains gnes, dont le gne BMP4
(bone morphogenetic protein) : plus intensment et
plus tt ce gne est exprim au cours du dvelop-
pement embryonnaire, plus le bec de ces oiseaux
devient robuste, ce qui a des consquences sur leur
alimentation. La protine BMP4est le produit de lex-
pressionde ce gne, elle agit sur ladiffrenciationdes
cellules lors du dveloppement
embryonnaire.
Si ungnehomotiquesexprime
un endroit inappropri, les
consquences sont visibles sur
le plan dorganisation. Chez les
serpents, le gne Hox6, respon-
sable de la formation de ctes,
sexprime tout le long de laxe
antropostrieur, contraire-
ment ce qui se passe chez les
autres vertbrs. Ceci explique
la prsence de ctes sur toutes
les vertbres des serpents.
Les variations de la localisation,
de la chronologie et de linten-
sit de lexpression des gnes
homotiques sont une source
importantedediversicationdes
tres vivants.
Des associations entre espces
diffrentes
Une diversicationdestresvivantsest possible sans
modication de leur gnome, par lassociation des-
pces diffrentes. Dans le cas des symbioses, les deux
individus tirent un avantage sassocier : changes
de substances nutritives, protection, hbergement,
dfensesont des exemples debnces rciproques.
La transmission de comportements
nouveaux
Chez les oiseaux, le chant est indispensable auchoix
des mles par les femelles pour la reproduction. Il
sagit dun comportement transmis par les parents
aux jeunes par apprentissage.
Chez les chimpanzs, le choix et lutilisation de
cailloux pour casser des noix sont transmis des
adultes aux jeunes de certaines communauts seu-
lement, durant un apprentissage de cinq annes.
Les comportements nouveaux sont transmis de g-
nration en gnration par voie non gntique.
Gospizes (10 20 cm) de gauche droite : fuligineux
bec pointu modeste des mangroves
Gnalogie du bl Gnalogie du bl
Engrainsauvage
Triticum urartu
Diplode
Aegilops speltoides
Diplode
Amidonnier
Anctredesblsdurs
T. turgidum
Ttraplode
T. tauschii = Ae. squarrosa
Diplode
Froment =bltendre
T. aestivum
Hxaplode=42chromosomes
peautre=blrustique
T. aestivum ssp spelta
Hxaplode
2n = 14
2n = 14
2n = 14
4n = 28
6n = 42
6n = 42
MOTS CLS
CELLULES
GERMINALES
Cellules lorigine des gamtes.
Leur patrimoine gntique est
transmissible la descendance de
lindividu.
CELLULES
SOMATIQUES
Touteslescellulesdunorganisme
lexceptiondes cellules germinales.
HYBRIDATION
Croisement de deux individus
appartenant deux espces diff-
rentes oudeuxvarits diffrentes
dune mme espce.
PLASMIDE
Chez les procaryotes, unplasmide
est une molcule dADN surnu-
mraire gnralement circulaire
pouvant tre transfre dautres
bactries.
PLODIE
Nombre de lots de chromosomes
que possde une cellule.
POLYPLODE
Se dit dunorganisme qui possde
plus de deux exemplaires de cha-
cun de ses chromosomes.
NOTION CL
LES GNES
HOMOTIQUES
Les gnes homotiques, ou ar-
chitectes, sont impliqus dans
lidentit cellulaire le long de laxe
antropostrieur lors du dvelop-
pement embryonnaire. Lexpres-
sion de chacun dentre eux est
restreinte un groupe de cellules
dlimit dans lespace, ce qui d-
termine le plan dorganisation.
Rpartis sur quatre chromosomes
chez les vertbrs et un seul chro-
mosome chez les insectes, ils for-
ment une famille multignique.
Les squences de ces gnes sont si
ressemblantes entreles diffrentes
espces quil peut tre possible,
par manipulation gntique, de
remplacer un gne homotique
dinsecte par son homologue de
vertbr, sans consquence sur le
phnotype.
NOTIONS CLS
EMPREINTE
Processus dapprentissage mis en
jeu au cours du dveloppement
des jeunes, et entranant une mo-
dication durable du comporte-
ment, qui peut tre hritable mais
de manire non gntique.
SYMBIOSE
Association bnfices rci-
proques entre deux organismes
appartenant des espces dif-
frentes.
Exemples de symbioses :
les lichens sont constitus
dune association entre une
algue unicellulaire et un cham-
pignon : lalgue fournit les pro-
duits de la photosynthse, tan-
dis que le champignon permet
la survie dans un milieu sec ;
certaines colonies de four-
mis sont en symbiose avec des
arbres tel le ccropia, elles y
sont hberges et nourries tout
en participant activement aux
dfenses de larbre contre les
herbivores ;
les mycorhizes sont une as-
sociation entre les racines des
vgtaux et des champignons ;
les nodosits des fabaces,
comme le haricot, associent les
racines de ces plantes des
bactries du genre Rhizobium :
la plante fournit les produits de
la photosynthse, tandis que les
bactries apportent les produits
azots issus de la fixation de
lazote atmosphrique.
Chimpanzs
DEUXARTICLESDUMONDECONSULTER
Des moules des quatre saisons en
France ? Lide nest pas sans danger p. 16
(J.-C. Rt, 21 novembre 2009)
Casser des noix, une question de culture
chez le chimpanz p. 17
(Jrme Grenche, 19 mai 2012)
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
LESSENTIEL DU COURS
14
UN SUJET PAS PAS
15 15 Gntique et volution Gntique et volution
Lintitul complet du sujet
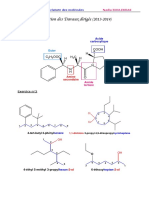
Chez les mammifres, le placenta constitue une zone
dchanges entre le sang maternel et le sang ftal,
ncessaire au dveloppement du ftus. En 2009,
des chercheurs ont mis lhypothse dune origine
virale de certains gnes indispensables la mise en
place du placenta.
Proposez les diffrentes tapes duscnario ayant pu
mener lapparition du placenta chez un anctre
des mammifres actuels an de montrer comment
les virus peuvent jouer un rle majeur dans la
diversication du vivant.
Votre rponse sappuiera sur lexploitation du dos-
sier. Aucune tude exhaustive des documents nest
attendue.
Document de rfrence
Le placenta, une zone d'changes entre le sang de
la mre et le sangde l'enfant qui ncessite la fusion
des cellules
Les documents
Document 1 : Un virus lorigine du placenta ?
Lapparition des mammifres placentaires, dont
lembryon est aliment et protg grce un pla-
centa, il y a prs de 100millions dannes, serait-elle
lie une contamination virale ? Cest lhypothse
que font des chercheurs aprs avoir dcouvert que
le gne syncytine A, dorigine virale, est essentiel
au dveloppement du placenta chez la souris. En
effet, les syncytines sont des protines de surface
permettant auxcellules qui les portent de fusionner
entre elles, comme le fait un virus pour entrer dans
une cellule. Ces fusions permettent la fabrication
dune nappe cellulaire multinucle*, constituant
essentiel du placenta, forme par la runion de
cellules individuelles en des syncytia . Pour les
chercheurs, qui ont publi leurs travaux en juin
2009, une contaminationvirale pourrait donc avoir
t un vnement fondateur dans le passage dun
dveloppement embryonnaire externe, chez les ani-
maux qui pondent des ufs, un mode interne .
* Multinucle : qui comporte de nombreux noyaux.
Source : daprs le journal du CNRS, n 236,
septembre 2009
Document 2 : Lecyclederplicationdunrtrovirus
Un rtrovirus est un virus dont le gnome est
constitu dARN. Sa particularit est de possder
une enzyme qui permet la transcription inverse
de lARNviral dugnome enmolcule dADN com-
plmentaire capable de sintgrer lADN de la
cellule-hte. Il utilise ensuite la machinerie cellulaire
pour se rpliquer.
Document 3 : Des rtrovirus endognes
Les rtrovirus infectieux possdent la proprit
remarquable de sintgrer dans lADN de nos chro-
Partie 2.2 : Lorigine
virale du placenta
partie
maternelle
cordon
ombilical
vaisseaux
sanguins
ftaux
P
L
A
C
E
N
T
A
sac amniotique
cordon
ombilical utrus
maternel
mosomes. En gnral, les cellules infectes sont des
cellules somatiques qui ne sont pas impliques dans
la transmission de notre patrimoine gntique.
Cependant, lorsquun rtrovirus parvient infecter
une cellule de la ligne germinale*, le rtrovirus
intgr peut se transmettre la descendance comme
nimporte quel gne : il devient alors un rtrovirus
endogne . Le gnome de tous les vertbrs est
ainsi envahi par de telles structures, et le squenage
systmatique dun grand nombre de gnomes, dont
ceux de lhomme et de la souris, montre que les
rtrovirus endognes reprsentent prs de 8 % du
matriel gntique de ces espces.
Fort heureusement, la plupart des rtrovirus endo-
gnes sont inactifs. Quelques rares lments sont ce-
pendant toujours capables de produire des protines
dorigine rtrovirale.
*Cellule de la ligne germinale : cellule lorigine de
gamtes.
Extrait dune publication du CNRS 2009
Source : http://www.cnrs.fr/insb/recherche/parutions/
articles09/t-heidmann.htm
Analyse du sujet
Il sagit de reconstituer une tape de lhistoire du
gnome des mammifres, dans laquelle les rtrovi-
rus infectieux semblent avoir jou un rle majeur.
Lapparition, par des mcanismes qui auront t
dduits des documents, dune structure nouvelle
comme le placenta est unexemple de diversication
gntique du vivant.
Pour le baccalaurat, il nest pas attendude connatre
tous les mcanismes de diversication de manire
dtaille. Ils sont tudier en relation avec leurs
consquences sur la diversicationdes tres vivants.
Les objectifs prioritaires sont la pratique du
raisonnement scientique et largumentation
partir de documents.
Le corrig
Le scnario ayant men lapparition du
placenta chez un anctre des mammifres
actuels :
Exploitation des documents 2 et 3
Infection par un rtrovirus dune cellule
germinale dun anctre des mammifres, par
fusion avec sa membrane.
Intgration de lADN viral, issu de la trans-
cription inverse de lARN viral, dans le
gnome de la cellule germinale.
Transmission la descendance via les ga-
mtes porteurs des gnes viraux.
Certains gnes viraux restent actifs chez les
descendants et sexpriment par la production de
protines virales.
Exploitation du document 1
Certaines protines virales de surface, les syncy-
tines, exprimes par les cellules des descendants,
permettent leur fusion pour former des syncytia.
Les gnes viraux codant pour ces protines de
fusion cellulaire sexpriment au moment du dve-
loppement embryonnaire et permettent la mise en
place dune zone dchange entre le sang de la mre
et le sang du ftus : le placenta.
Les virus joueraient un rle majeur dans la
diversication du vivant
Le squenage des gnomes de vertbrs montre
que les gnes issus de rtrovirus reprsentent prs
de 8 % du matriel gntique.
Dans lexemple des syncytines, lintgration et
lexpression de gnes viraux a permis lapparition
du placenta.
Cette structure nouvelle a permis de passer dun
dveloppement dans des ufs undveloppement
interne.
AUTRES SUJETS POSSIBLES AU BAC
SUR CE THME
Partie 2.1 :
Exploiter des documents pour tablir le lien
entre des variations de lexpression de gnes ho-
motiques et un exemple de diversication du
vivant.
Exploiter des documents sur lvolution du g-
nome des espces cultives.
Partie 2.2 :
Exploiter des documents pour montrer que cer-
tains exemples de diversicationduvivant se ra-
lisent sans modicationdugnome.
ZOOM SUR
LE CYCLE DE RPLICA-
TION DUN RTROVIRUS
Un rtrovirus est constitu dARN
dans une enveloppe protique.
Il peut infecter les cellules qui
prsentent leur surface des pro-
tines auxquelles il peut samar-
rer : ce sont ses cellules-cibles. Le
virus peut alors y transfrer son
matriel gntique enfusionnant
avec lamembranecellulaire. LARN
viral est converti enADNpar rtro-
transcription grce une enzyme
virale : la transcriptase inverse.
LADNviral sintgreaugnomede
la cellule-hte et sexprime. Ainsi,
ARN viraux et protines virales
sont produits, permettant las-
semblage de nouvelles particules
virales qui vont tre dissmines.
DES VIRUS PARTOUT !
Ltude des milieux naturels
partir des gnomes qui sy trou-
vent, la mtagnomique, permet
dsormais le comptage et lidenti-
cationdes virus de latmosphre,
malgrleur petitetaille, infrieure
au micromtre. Rsultat : dans un
mtre cube dair, on trouve entre
1,7 et 40 millions de virus selon
les saisons. Pour les bactries, la
fourchette est plus basse : entre
860 000 et 11 millions dindivi-
dus par mtre cube. Au repos, un
adultepompeenmoyenne10litres
dair par minute. chaque minute
qui passe, entre17 000et 400000
viruspntrent ainsi dansnospou-
mons De quoi pousser un hypo-
condriaque cesser de respirer !
Latmosphre est un rservoir de
virus encore largement inexplor
auquel il serait temps de sintres-
ser, notamment pour identier des
entits susceptibles de sattaquer
aux cultures... et aux hommes.
Source : daprs Pierre Barthlmy,
Combien de virus inhalez-vous chaque
minute ? sur le blog Passeur de
sciences, le 10 octobre 2012
ZOOM SUR...
LENDOSYMBIOSE
Lendosymbiose est une coopra-
tionbncesmutuelsentredeux
organismes, lun tant contenu
dans lautre. Les cellules eucaryotes
possdent des mitochondries, des
organites cellulaires spcialiss
dans la production dnergie qui
contiennent de lADN. Une com-
paraison gntique a permis de
montrer de grandes similitudes
entrelegnomedesmitochondries
et celui des bactries de type Ric-
kettsia, ce qui suggre une origine
endosymbiotique de ces organites
cellulaires. Toutefoislegnomemi-
tochondrial aperducertains gnes,
qui sont inclus dans le gnome du
noyaude la cellule eucaryote.
UNE LIMACE MARINE
CHLOROPHYLLIENNE
La limace marine Elysia chlorotica
est marronsanaissanceet broute
des algues. Elle devient alors verte.
la lumire dusoleil, la limace d-
ploie des expansions de son corps
des parapodes pour capter la
lumire. leve lobscurit, elle
se replie, redevient marron et doit
brouterdesalgues. lalumire, elle
na pas besoinde manger car lani-
mal agit commeunvgtal chloro-
phyllien, produisant les molcules
dont elle a besoin grce la pho-
tosynthse. Des chloroplastes ont
t extraits des premires algues
manges. Ils sont conservs dans
les cellules de la paroi intestinale
mais ne sont pas transmis la des-
cendance. Des gnes ncessaires
la photosynthse se trouvent dans
le gnome de la limace. Ils ont d
tre acquis au cours de lvolution
par transfert horizontal entre les
algues et une anctre de cette li-
mace. Cetransfert apermis lamise
en place de cette endosymbiose
particulire.
Elysia chlorotica (2 3 cm)
Ce quil ne faut pas faire
Recopier lidentique des phrases issues des
documents.
Utiliser systmatiquement les documents dans
lordre du sujet.
tudier les documents de manire exhaustive
sans les mettre en relation.
LESSENTIEL DU COURS UN SUJET PAS PAS
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
16
LES ARTICLES DU
17
LES ARTICLES DU
17 Gntique et volution Gntique et volution
Des moules des quatre
saisons en France ?
Lide nest pas sans danger
LIfremer vient de dposer un brevet pour dvelopper les recherches sur les moules
triplodes, linstar des hutres, minimisant les risques de ces exprimentations pour le
milieu naturel.
A
prs les hutres, bientt
des moules triplodes
dans nos assiettes ? Se-
lonnos informations, lInstitut
franais de recherche pour lex-
ploitation de la mer (Ifremer)
vient de dposer unbrevet inti-
tul Obtentionde mollusques
bivalves ttraplodes partir de
gniteurs diplodes . Derrire
cet nonc barbare se cache la
poursuite des investigations
dans le but de caractriser
les performances biologiques
des moules triplodes dites
chimiques, obtenues via une
induction chimique partir
de gniteurs diplodes, et des
moules triplodes dites natu-
relles, issues ducroisement de
femelles diplodes et de mles
ttraplodes .
En clair, il sagit de crer une
moule des quatre saisons ,
produite et vendue toute lan-
ne (aujourdhui, la priode
de maturation/ponte stale de
fvrier mai) an den limiter
limportation (40 000 tonnes
par an).
Le problme est que les tri-
plodes ne sont pas sans
danger. Les ostriculteurs en
savent quelque chose. Cet t
encore, des bassins dhutres
ont t frapps de mortalits
considrables, parfois jusqu
80 %. Certains ostriculteurs
tiennent, non sans raison,
pour responsables les hutres
triplodes, apparues sur le
march depuis quelques an-
nes. Ces hutres, obtenues
la suite dune manipulation
chimique mise au point, en
1997, par lIfremer, sont rpu-
tes striles. Elles sont cen-
ses ne pas se reproduire. La
dure de leur croissance est
infrieure de 40 % : produites
en deux annes, leur rentabi-
lit est bien meilleure. Seule
ombre au tableau, la manipu-
lation ou castration chimique
ne peut tre effectue que
dans quelques laboratoires
closeries qui disposent
dsormais dun monopole de
production. Une hutre sur
deux, aujourdhui, serait une
triplode !
LIfremer a d admettre, cet
t, que certaines triplodes
ntaient pas entirement st-
riles et se reproduisaient en
milieu naturel. Nos scienti-
ques seront-ils les derniers
reconnatre les ravages
gntiques des polyplodes
cres par les contaminations
des triplodes sur les stocks
naturels de gigas [hutres
creuses] ? Regardez donc les
noyaux des cellules, cest le
bordel l-dedans ! , proteste
un ostriculteur sur le site
Internet Ostrea.com.
La loi du silence
Les triplodes seraient donc
lorigine de la mortalit des
hutres juvniles. Beaucoup le
pensent, peu le disent. Indirec-
tement, cest lIfremer qui est
en cause. Indirectement, car
cet organisme public (EPIC),
cr en 1984, est la fois le
mdecin et la police des mers.
Seule lIrlande a rompu la loi
du silence et interdit jusqu
nouvel ordre limportation
de naissains en provenance
des closeries franaises pour
protger ses bassins. Certains
professionnels ont dcid de
prendre leur destin en main.
Sous lemblme Ostricul-
teur traditionnel , une tren-
taine de professionnels de sept
bassins diffrents entendent
valoriser les hutres nes en
mer.
Le consommateur doit avoir
le choix entre consommer une
hutre issue du milieu naturel,
leve en trois ans, et une
hutre strile issue de labora-
toire, leve en deux ans ,
prcise Laurence Maho, du
rseau Cohrence. Dans
quelques annes, elle devra
peut-tre ajouter les moules
dans son combat.
J.-C. Rt
(21 novembre 2009)
POURQUOI
CET ARTICLE ?
Cet article paru dans la ru-
brique gastronomie montre
que la polyplodisation, qui est
un processus naturel, peut-
tre provoque chez certaines
espces consommes par
lhomme, an den tendre la
priode de production. Cette
pratique ne semble pas sans
danger et pourrait augmenter
la mortalit des mollusques,
comme lont expriment les os-
triculteurs. De plus, contraire-
ment ce qui tait prvu, les tri-
plodes pourraient se reproduire
et contaminer les autres
hutres. Cet exemple montre
les tentatives de lhomme pour
matriser la nature des ns de
consommation, mais aussi les
limites de ces pratiques et la
ncessit de connaissances ap-
profondies sur les processus de
diversicationdes tres vivants.
Casser des noix, une question
de culture chez le chimpanz
La forme et la matire des outils utiliss varient selon la communaut dappartenance
des primates.
L
a culture est-elle vri-
tablement le propre de
lhomme ? La questionest
relance par une tude sur le
chimpanz paratre, le 22 mai,
dans la revue Current Biology
et conduite par Lydia Luncz,
de lInstitut Max-Planck dan-
thropologie volutionniste de
Leipzig. Pendant neuf mois, la
primatologue a tudi les com-
portements de slection dou-
tils destins casser des noix
de coula chez les chimpanzs
sauvages (Pan troglodytes
verus) de trois communauts
voisines de la fort de Ta, en
Cte dIvoire.
Lutilisation doutils pour cas-
ser ces noix, trs nourrissantes
mais presque aussi dures que
de la pierre, est certainement
le comportement le plus so-
phistiqu observ chez ces
primates connus pour leur
organisationsociale complexe.
Tous ont la mme technique,
qui consiste placer le fruit
sur une racine qui sert den-
clume et le frapper laide
dun percuteur en bois ou en
pierre. En tenant compte de
la disponibilit des outils de
cassage et de la rsistance des
noix de coula au cours de la
saison, ltude rvle des diff-
rences dans le type et la taille
des outils slectionns entre
les communauts riveraines
de chimpanzs.
Endbut de saison, ces groupes
choisissent principalement des
pierres, plus dures que le bois,
pour casser les noix encore
fraches. Mais, la saison avan-
ant, quand les noix devien-
nent sches donc plus faciles
casser, les communauts du
nord et de lest de la fort de
Ta privilgient les marteaux
en bois, plus abondants. Seuls
les chimpanzs situs au sud
continuent dutiliser des mar-
teaux en pierre pendant toute
la saison.
La culture est avant tout un
phnomne social. Les com-
munauts de chimpanzs qui
vivent dans le mme environ-
nement gardent nanmoins
leurs prfrences pour des
types doutils de cassage dif-
frents , souligne Christophe
Boesch, coauteur de ltude
et directeur du dpartement
de primatologie de lInstitut
Max-Planck. Il y a transmission
dune tradition entre gnra-
tions au sein dun groupe.
Adaptation des
comportements
Selon les communauts, les
chimpanzs slectionnent
aussi des marteaux en bois de
tailles variables : petits aunord
et grands dans les deux autres
groupes. Les diffrences
culturelles dans la slection
des outils sont maintenues
malgr quelques changes
dindividus entre les commu-
nauts de chimpanzs, notam-
ment lors de la migration des
femelles dune communaut
lautre , ajoute le chercheur.
Des groupes riverains auraient
ainsi dvelopp leur propre
cultureandadapter leur com-
portement de cassage selon la
rsistance des noix pendant la
saison.
Jusqu prsent, les primato-
logues pensaient que seuls les
chimpanzs vivant dans des
territoires gographiquement
loigns utilisaient des outils
diffrents pour casser les noix
de coula. Les populations tu-
dies par le pass prsentent
beaucoup de diffrences dans
lusage doutils, mais largu-
ment des diffrences cultu-
relles a t mis en doute, no-
tamment enraisondepossibles
variations dordre cologique
ou gntique. En effet, 100
1 000 km sparent les popula-
tions de chimpanzs qui utili-
sent des outils diffrents pour
se nourrir, prcise Christophe
Boesch. Cette fois-ci, notre
tude souligne le maintien de
ces diffrences culturelles chez
des chimpanzs vivant dans
des environnements similaires
et avec un certain brassage g-
ntique, ce qui est difcilement
contestable. Reste savoir si
ces diffrences sont lies des
diffrences defcacit , cone
le chercheur.
Longtemps sujet contro-
verse, la question de lhritage
culturel chez le chimpanz
resurgit donc avec ltude de
Lydia Luncz, qui met en vi-
dence de fortes similarits avec
lhomme. Chez les humains,
les diffrences culturelles sont
une partie essentielle de ce qui
distingue les groupes voisins
qui vivent dans des environ-
nements trs similaires. Pour
la premire fois, une situation
trs semblable a t observe
chez les chimpanzs sauvages
vivant dans le parc national de
Ta, et cela dmontre quils par-
tagent avec nous une capacit
de diffrenciation culturelle
une chelle ne , conclut
Lydia Luncz.
Ces travaux qui rvlent la di-
mensionculturelle ducompor-
tement des chimpanzs souli-
gnent lurgente ncessit de
prserver lhabitat de cette es-
pce menace dextinction.
Pour son bien propre mais
aussi pour le ntre : la dispari-
tion de ces hominids qui par-
tagent plus de 98 % de leurs
gnes avec Homosapiens pour-
rait compromettre la compr-
hension des racines de la
culture humaine.
Jrme Grenche
(19 mai 2012)
POURQUOI
CET ARTICLE ?
Cet article rvle que la tech-
nique du cassage des noix
de coula par les chimpanzs
est plus complexe quil ny
parat et quil sagit dune
question de culture . Ce
comportement transmis
par apprentissage est un
exemple de diversification
non gntique des tres
vivants. La question de la
culture animale reste sujette
controverse tant elle fait
vaciller les certitudes quant
aux spcificits de lespce
humaine. En sciences de la
vie et de la terre, lhomme
doit tre tudi comme toute
autre espce.
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
18
LESSENTIEL DU COURS
19
LESSENTIEL DU COURS
De la diversication des
tres vivants lvolution
de la biodiversit
La diversit des populations change
au cours des gnrations
Dans une population, la frquence des diffrents
allles peut voluer de manire alatoire si ces allles
napportent ni avantageni dsavantageauxindividus
qui les portent. Le hasard des brassages au cours de
la reproduction sexue dtermine lvolution de
la population, de faon dautant plus marque que
celle-ci est petite. Il sagit de la drive gntique.
Dans un milieu donn, selon les allles dont ils sont
porteurs, certains individus vont mieux survivre (ils
senourrissent plus facilement ouchappent auxpr-
dateurs) et/ ou davantage se reproduire (ils attirent
davantage les partenaires). Ces individus engendrent
plus de descendants que les autres et transmettent
ainsi davantage leurs allles aux gnrations sui-
vantes. La consquence dans la population est une
augmentation de la frquence des allles dont ces
individus sont porteurs, cest la slection naturelle.
Lespce : un concept dlicat dnir
La dnition de lespce a t modie au cours de
lhistoire de la biologie. Plusieurs dnitions se sont
succd et reposent sur des critres varis.
Cuvier (1769-1832) : Une espce est un ensemble
dtres vivants partageant des critres anatomiques
et physiologiques. Or, deuxanimauxseressemblant
fortement peuvent ne pas appartenir la mme
espce, car ils ne rpondent pas la dnition biolo-
gique de lespce.
Dnition biologique adopte par Buffon (1707-
1788) : Une espce est une communaut dtres vi-
vants pouvant produire des descendants eux-mmes
fconds (critre dinterfcondit).
Dnition cologique : une espce est une popu-
lation adapte une niche cologique particulire.
Dnition gntique : Deux individus doivent
avoir le mme nombre de chromosomes et sur
chaquechromosomelemmenombredenuclotides
pour appartenir la mme espce Dawkins (1941-).
Actuellement, on considre que les espces sont
des populations dindividus sufsamment isoles
gntiquement des autres populations .
Dans tous les cas, uneespcenest dniequedurant
uncertainlaps detemps, dlimitpar sonapparition
(spciation) et son extinction ou son volution vers
une autre espce.
La spciation ou cration
de nouvelles espces
Plusieurs mcanismes permettent dexpliquer la
naissancedunenouvelleespcepartir duneespce
ancestrale.
D
epuis lapparition de la vie sur la Terre et leur hypothtique
anctre commun LUCA (last universal common ancestor),
les tres vivants se sont diversis. La biodiversit, qui
se dnit diffrentes chelles, correspond notamment la di-
versit des espces, mais aussi la diversit au sein de celles-
ci. Au cours de lhistoire de la Terre, la biodiversit a explos
certaines priodes et a chut lors de crises biologiques, en lien
avec des modications des milieux de vie. Ainsi, la biodiversit
volue constamment selon divers mcanismes, les innovations
se maintenant ou non dans les populations sous leffet de la d-
rive gntique et de la slection naturelle.
temps
spciation
uneespce
unindividu
Une vision thorique de lespce
extinction
spciation
uncroisement
spciation
uneespce
(DaprsGuide critique de lvolution G. Lecointre d. Belin 2009)
Un rseau gnalogique fictif (cercles = individus, traits = croisements)
La sparation de populations par un obstacle
physique. En effet, la spciation peut tre lie la
sparationde populations par une barrire physique
(cours deau, relief) qui va empcher les changes g-
ntiques entre deuxpopulations de la mme espce
initiale et aboutir la divergence en deux espces.
La colonisationdune nouvelle niche cologique.
Des individus fondateurs se sparent de lespce
mre pour coloniser une nouvelle niche, isole (le)
ou adjacente, mais soumise dautres pressions
de slection. Comme peu dindividus ont migr,
tous les allles de la population de dpart ne sont
pas forcment reprsents aux mmes frquences
que dans la population dorigine. Les facteurs en-
vironnementaux pouvant tre diffrents dans le
nouveau milieu colonis, la pression de slection
peut favoriser certains allles ou le maintien de
certaines innovations gntiques.
La spciation sans isolement gographique.
Dans un mme milieu, au sein dune population,
lapparitiondunnouvel allle peut aboutir former
une nouvelle espce qui exploite diffremment les
ressources de lenvironnement ou ne peut plus se
reproduire avec lespce initiale. En cause : lincom-
patibilit des parades sexuelles ou, pour des oiseaux
ou des grenouilles, le fait que leurs chants soient
devenus trs diffrents et ne leur permettent plus
de se reconnatre.
Lindividualisation dune nouvelle espce peut
prendre une dure variable, de quelques annes
des millions, selon la population de dpart, lespce
considre, le milieu et les possibilits dchanges
gntiques entre les individus.
Lextinction des espces
Une espce est considre comme teinte si len-
semble de ses individus a disparu ou sils cessent
dtre isols gntiquement. Il existe des extinctions
locales (dans une niche cologique donne) ou
globales ( la surface de la Terre).
Lextinctiondune espce peut entraner celle dune
autre espce. Ainsi, si les abeilles disparaissent,
toutes les espces vgtales dont la pollinisation
est strictement dpendante de ces insectes dispa-
ratront.
Au cours de lhistoire de la Terre, des diminutions
massives de la biodiversit ont eu lieu : la priode
crtac-tertiaire est unexemple de crise biologique,
avec notamment la disparition des ammonites et
des dinosaures (dont les oiseaux sont les descen-
dants).
Population
dedpart
Initiation
delaspciation
Accroissement
delisolement
reproductif
Nouvelles
espces
Dansuneniche
isole
Colonisation
dunenouvelle
niche
lintrieur dune
population
Polymorphisme
gntique
Par sparation par
un obstacle physique
Par colonisation
dune nouvelle niche cologique
Sans isolement
gographique
TROISARTICLESDUMONDECONSULTER
Ltrange vie si peu sexuelle de la petite
fourmi de feu p. 22
(Stphane Foucart, 2 juillet 2005)
La naissance des espces, un processus
surprises p. 22-23
(Stphane Foucart, 23 mai 2006)
Lvolution lenvers p. 23
(Nicolas Gompel, Benjamin Prudhomme,
12 mai 2012)
ZOOM SUR
MME ESPCE OU
ESPCES DIFFRENTES ?
Les lions (Panthera leo) et les
tigres (Panthera tigris) peuvent
engendrer des descendants uni-
quement en captivit, mais leurs
petits (tigrons, ligres) sont striles.
Ils nappartiennent donc pas la
mme espce
Lefuligulemorillon(Aythyafuligu-
la) et lefuligulemilouin(Aythyafe-
rina) sont des canards qui peuvent
donner exprimentalement des
descendants fertiles alors quils
nhabitent pas les mmes milieux.
Ils sont considrs comme deux
espces diffrentes.
LES PINSONS
DES GALPAGOS
Chezles pinsons deDarwin, qui oc-
cupent diffrentes les, des formes
de bec varies ont t slection-
nes en fonction de la nourriture
disponible.
Une population dindividus issus
dune espce A colonise une le.
Au bout dun certain temps, elle
peut devenir une espce B, dont
certains individus colonisent une
autre le et forment une espce C.
Cette dernire colonise les les
alentours, dont celle qui est dj
occupe par lespce B. Elles ne
sont pas interfcondes. Leur pos-
sibilit de cohabitation dpend
du fait quelles occupent ou non
la mme niche cologique. Si cest
le cas, les deuxespces entrent en
comptitionce qui peut entraner
la disparition de lune delles.
ZOOM SUR...
Des exemples de spciation
Dans larchipel de Madre, six
espces apparentes aux souris
domestiques europennes pr-
sentent entre 22 et 30 chromo-
somes au lieu de 40, suite la
fusionde certains chromosomes.
Ces populations non interf-
condes proviennent de lvolu-
tion de souris apportes par les
bateaux des dcouvreurs de cette
le au relief escarp (les Vikings
au IX
e
sicle, les Portugais au
XV
e
sicle). Il sagit dun exemple
de spciation par isolement go-
graphique.
Dans les sous-sols de Londres,
des moustiques Culex pipiens
molestus sont gntiquement
diffrencis selon les lignes de
mtro. Ils ont volu depuis une
centaine dannes partir dune
espce de surface, avec laquelle
ils ne sont plus interfconds. La
profusion de leurs proies (les
mammifres, humains comme
rongeurs) transitant par ce
moyen de transport, la chaleur
des lieuxet la prsence de aques
deau ont acclr le rythme de
reproduction (plusieurs cycles
par an), ce qui a permis une sp-
ciation rapide.
Aux tats-Unis, les larves des
mouches Rhagoletis consom-
ment les baies de laubpine. Le
dveloppement de la culture des
pommes a favoris lindividua-
lisation, sans isolement gogra-
phique, dune nouvelle espce
consommatrice de pommes.
Gntique et volution Gntique et volution
Des mcanismes de spciation
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
LESSENTIEL DU COURS
20
UN SUJET PAS PAS
21 21
Lintitul complet du sujet
Il existe actuellement deux espces de pandas : le
panda roux (Ailurus fulgens) et le grand panda (Ai-
luropoda melanoleuca). Tous deux sont vgtariens,
se nourrissant de grandes quantits de feuilles de
bambou. Ils prsentent par ailleurs une particularit
anatomique remarquable : la prsence dun sixime
doigt (ou faux pouce ) chaque main, longtemps
interprte comme une adaptation leur rgime
alimentaire.
En2005, ona dcouvert enEspagne unfossile dat de
9millionsdannes(Simocyonbatalleri) apparentaux
pandasactuelset prsentant lui aussi unsiximedoigt.
Montrez comment linterprtation du sixime doigt
des pandas entermes dadaptationaurgimealimen-
tairevgtarien(adaptationdont vous expliquerezles
mcanismes) sest nuance la lumire de nouvelles
dcouvertes.
Votre rponse sappuiera sur lexploitation du dos-
sier. Aucune tude exhaustive des documents nest
attendue.
Document de rfrence : Les trois pandas connus
Les documents
Document 1 : Reprsentationactualisedes relations
de parent entre ours et panda
Document 2
Les grands pandas gants sont des ours duntype bien
dni, membres de lordre des carnivores. Les ours
ordinairessont lesreprsentantslesplusomnivoresde
leur ordre, mais les pandas ont restreint luniversalit
deleursgots: ilsdmentent lappellationdeleurordre
en tirant leur subsistance presque exclusivement du
bambou. [] Assis biendroit sur leur derrire, ils mani-
pulent sestigesavecleurspattesavant, sedbarrassant
des feuilles pour ne consommer que les pousses. []
Commentledescendantduneligneadaptelacourse
peut utiliser ses mains de faon si habile ? Ils tiennent
lestigesdebamboudansleurspatteslesdpouillent de
leurs feuilles enfaisant passer les tiges entre unpouce
apparemment exibles et les autres doigts. []
[] Anatomiquement, le pouce du panda
nest pas un doigt. Il est construit partir dun os
appel le ssamode radial (du radius), normale-
ment un des petits os formant le poignet. Chez le
panda, le ssamode radial est trs dvelopp et
si allong que sa taille atteint presque celle des os
des phalanges des vrais doigts. [] Lallongement
du ssamode radial a pu tre provoqu par une
transformation gntique, peut-tre une seule
mutation affectant le rythme et la vitesse de
la croissance. [] Le vrai pouce du panda, trop
spcialis pour tre utilis une autre fonction
et devenir un doigt opposable,
apte la manipulation, est re-
lgu un autre rle. Le panda
est donc contraint de se servir
des organes disponibles et de
choisir cet os du poignet hy-
pertrophi, solution quelque
peu btarde mais trs fonc-
tionnelle.
Le faux pouce du panda gant
est en ralit un os du carpe
Partie 2.2 : Le sixime
doigt des pandas
Simocyon
Ailurus
Ailuropoda
Middle Middle Late Late Late Early Early
40 30 25 20 15 10 5 0 Mya. 35
Autres Ours
(os plat de la paume)
transform en pouce
opposable. (Not rs pour
os ssamode radial)
Source : Extrait de Le Pouce
du panda ou les grandes
nigmes de lvolution,
Stephen Jay Gould, 1980
Document 3
En tudiant sa den-
ture, nous sommes arrivs la conclusion que cet
animal mangeait essentiellement de la viande, et
non des vgtaux comme le petit panda actuel,
annonce Stphane Peign, jeune chercheur au
Laboratoire de gobiologie, biochronologie et
palontologie humaine. Cest pourquoi nous pen-
sions que Simocyon nutilisait pas son sixime
doigt pour saisir les pousses de bambou comme
le fait aujourdhui le petit panda, mais plus cer-
tainement pour aider sa locomotion dans les
arbres. Et comme les donnes recueillies sur le
site indiquent quil vivait dans un environnement
peupl de nombreux prdateurs, cette trange
factie de lvolution de doter Simocyon dun faux
pouce apparat, dans ce contexte, vitale pour ce
carnivore plutt charognard et peu vloce : il
pouvait donc leur chapper en grimpant aisment
dans les arbres , poursuit le palontologue
Source : extrait dun communiqu de presse du
CNRS (Centre national de la recherche scientifique)
propos de la dcouverte de Simocyon batalleri
(mars 2005)
Analyse du sujet
La rponse argumente doit sappuyer sur les
informations pertinentes des documents, mises
en relation entre elles et avec les connaissances.
Elle comportera deux parties distinctes : une
proposition de scnario ayant conduit lappa-
rition et au maintien du sixime doigt chez les
pandas (mcanismes volutifs) ; puis comment
linterprtation de cette adaptation a t nuance
suite la dcouverte dun fossile.
Proposition de corrig
Diffrents scnarios volutifs peuvent tre accep-
ts sils sont plausibles et sans contradiction avec
les documents.
I. Mcanismes ayant conduit lapparition et
au maintien du sixime doigt chez les pandas :
proposition de scnario
Exploitation du document 1
Les pandas ont des liens de parent avec les ours
(plus particulirement le grand panda) et lespce
fossile Simocyon est un parent plus proche des
pandas roux que du grand panda.
Exploitation du document 2
Les pandas actuels ont un sixime doigt adapt
leur rgime alimentaire de type vgtarien
(bambou). Cette particularit anatomique a peut-
tre une origine gntique : une mutation dans un
gne qui modifie la croissance de los ssamode
radial.
Cette innovation gntique serait due au hasard,
faisant apparatre le faux pouce probablement
chez lanctre commun tous les pandas.
Certaines innovations peuvent donner un avan-
tage aux individus dans un milieu donn. Lin-
novation a dans ce cas plus de chances dtre
transmise la descendance et de diffuser dans la
population : cest la slection naturelle.
Il peut donc y avoir conservation de linnovation
si elle nest pas une gne et diffusion si elle
confre un avantage slectif, dans deux situations
diffrentes : chez Simocyon carnivore pour fuir les
prdateurs (document 3), chez les pandas pour se
nourrir (document 2). Cette innovation na pas t
conserve dans le groupe des ours.
II. Comment linterprtation du sixime doigt
des pandas a t nuance suite la dcouverte
dun fossile
Avant la dcouverte : le sixime doigt des pandas
tait interprt comme une adaptation leur
rgime alimentaire.
Aprs la dcouverte de Simocyon : il est interprt
comme une adaptation chez Simocyon pour fuir
les prdateurs en grimpant aux arbres ; chez les
pandas, pour saisir les branches de bambou.
Linnovation, partage par les pandas gants, les
pandas roux et Simocyon serait apparue chez lun
de leurs anctres communs, mais naurait pas t
conserve chez les ours.
Osdelamaindupandagant
Ce quil ne faut pas faire
Faire une analyse exhaustive de tous les docu-
ments sans lien avec la problmatique.
Se dispenser dapporter des connaissances.
Omettre dindiquer clairement les relations entre
les diffrents arguments.
AUTRES SUJETS POSSIBLES AU BAC
SUR CE THME
Partie 1 :
Expliquer les mcanismes qui dterminent
lvolution de la frquence dun allle dans une
population(drive gntique, slectionnaturelle).
Partie 2. 1 :
Nuancer la dnition de lespce partir
dexemples dhybrides interspciques.
Partie 2. 2:
Analyser, partir de documents, des exemples
de spciation dans des contextes et selon des m-
canismes varis.
ZOOM SUR
La vie de Charles Darwin
1809
Le 12 fvrier : naissance Shrews-
bury (Angleterre) de Charles
Darwin. En France, Lamarck pr-
sente sa thorie transformiste.
1831
Le 27 dcembre, Darwin sem-
barque comme naturaliste sur le
Beagle pour un voyage autour du
monde qui durera cinq ans.
1835
Le Beagle fait escale aux Gal-
pagos, o Darwin note des va-
riations dans la forme des becs
des pinsons, ce qui inspirera sa
thorie.
1837
Premire esquisse, par Darwin,
dun arbre volutionnaire gu-
rant dans sonNotebook onTrans-
mutation of Species.
1858
Prsentation Londres darticles
en commun avec Wallace sur la
perptuation des varits et des
espces par les moyens naturels
de la slection.
1859
Publication de De lorigine des
espces par voie de slection
naturelle ou la Prservation des
races favorises dans la lutte pour
la vie.
1871
Darwinlivre ses vues sur lorigine
de lhomme dans La Filiation de
l homme et la slection lie au
sexe.
1882
Mort de Darwin dans sa demeure
de Down, dans le Kent. Il sera en-
terr labbaye de Westminster.
Source: DatesclsdelaviedeCharles
Darwin , Le Monde, 6 fvrier 2009.
PERSONNAGE
CL
STEPHEN JAY GOULD
(1941-2002)
Palontologue amricain, coau-
teur de plusieurs ides tho-
riques sur lvolution. Il doit
sa renomme ses nombreux
essais, dont Le Pouce du panda,
et sa participation active la
lutte contre le crationnisme aux
tats-Unis.
ZOOM SUR...
LA PHALNE DU BOULEAU
Papillon nocturne qui existe sous
une forme claire, camoue des
prdateurs sur les troncs clairs du
bouleau, et une forme sombre.
Ltude de lvolution de la fr-
quencerelativedeces deuxformes
au sein des populations de pa-
pillons partir du XIX
e
sicle en
Angleterre constitue un exemple
classique pour expliquer le prin-
cipe de la slection naturelle. La
forme sombre, observe pour la
premire fois en 1848, est deve-
nue majoritaire dans les rgions
industrialises, o les troncs des
arbres deviennent plus sombres.
La survie diffrentielle des deux
formes de papillons, qui chap-
pent plus ou moins facilement
leurs prdateurs sur les troncs
clairs ou sombres, dtermine le
nombre de leurs descendants et
lvolution de la frquence des
deux allles impliqus.
SPCIATION ET
SLECTION NATURELLE
Aux tats-Unis, les valles de
Sacramento et San Joaquim sont
trop sches pour permettre
la salamandre de Californie dy
vivre. Encolonisant le milieu, une
partie de la population initiale a
contourn les valles sches par la
zone montagneuse de lest, tandis
que lautre est passe par la zone
ctire louest. Les salamandres
des zones montagneuses sont ta-
chetes, ce qui leur assure un bon
camouage vis--vis des prda-
teurs. Les salamandres des zones
ctires sont rouges, comme les
animaux venimeux vivant dans
ce milieu. Les deux populations
ont poursuivi leur migration vers
le sud jusqu la convergence des
deux valles : les salamandres de
lest et de louest ny sont plus
interfcondes, il sagit de deux
espces diffrentes.
Gntique et volution Gntique et volution
LESSENTIEL DU COURS UN SUJET PAS PAS
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
22
LES ARTICLES DU
23
LES ARTICLES DU
23
Ltrange vie si peu sexuelle
de la petite fourmi de feu
L
a brouille est ancienne.
Elle dure, sans doute, de-
puis plusieurs centaines
de milliers dannes. Depuis
suffisamment longtemps, en
tout cas, pour que les mles et
les femelles reproductrices de
Wasmannia auropunctata ne
partagent aujourdhui presque
plus rien. Pas mmeles parts les
plus tnues de leur organisme :
les gnomes des mles et des
reines decetteespcedefourmi
invasive sont si diffrents que
les biologistes pourraient croire
deux espces distinctes.
Comment celle que lonappelle
la petite fourmi de feu en
est-elle arrive une telle
extrmit ? Les chercheurs
franais et suisses qui ont mis
au jour ce miracle de lvolu-
tion expliquent, jeudi 30 juin
dans la revue Nature, ltrange
stratgie reproductive de Was-
mannia auropunctata. Dans ce
mode de reproduction jamais
observ auparavant, les mles
sont les clones de leur pre et
les reines sont les clones de
leur mre.
Chez la plupart des espces de
fourmis, les reines fabriquent
deuxtypes dufs, expliqueDe-
nis Fournier, premier auteur de
ltude, charg de recherche au
Fonds national de la recherche
scientique (FNRS) de Belgique.
Certains, dits haplodes, non
fconds, produisent des
mles tandis que dautres, dits
diplodes, sont fconds et
produisent soit des reines, soit
des ouvrires. Mais chez la
petite fourmi de feu, les reines
ont, peu peu, appris se pas-
ser de la semence des mles
pour engendrer dautres reines,
transmettant ainsi la totalit de
leur patrimoine gntique la
gnrationsuivantedefemelles
reproductrices.
Ducoup, les mles netransmet-
tent plus leurs gnes quaux
ouvrires, qui, striles, ne peu-
vent assurer la prennit de
ce patrimoine gntique. Pour
contrecarrer cet hgmo-
nisme gntique des reines,
les mles ont d ruser. Nous
pensons, sans en tre certains,
que les mles fcondent les
ovules haplodes et quune fois
la fcondation effectue, sous
certaines conditions, lADN du
mle peut liminer celui de la
femelle , prcise M. Fournier.
Luf en question engendre,
en dnitive, un clone du mle
layant fcond. Cest une
manire de parasitisme g-
ntique, selon M. Fournier. Le
mle utilise en quelque sorte
la reine comme une mre por-
teuse.
Chez Wasmannia auropunc-
tata, tout ou presque se passe
donc comme si les mles et les
femelles appartenaient deux
espces diffrentes. Cepen-
dant, mme si ce divorce,
consomm de longue date, les
place sur deux branches dis-
tinctes de larbre de lvolu-
tion, mles et reines ont mal-
gr tout besoin lun de lautre.
Leurs gnes se mlent ainsi
pour engendrer les ouvrires.
Bien que striles, ces dernires
nenassurent pas moins lorga-
nisation sociale de lespce et
le bon fonctionnement des
colonies.
Stphane Foucart
(2 juillet 2005)
POURQUOI
CET ARTICLE ?
Lexemple de la petite
fourmi de feu est pertinent
pour aborder la complexit
de la dnition de lespce.
Les stratgies reproductives
de cet insecte ont conduit
des volutions distinctes des
mles et des reines. Tout se
passe ainsi comme si les mles
et les femelles, bien quas-
socis au sein des colonies,
appartenaient des espces
diffrentes.
La naissance des espces,
un processus surprises
O
n sait comment et pour-
quoi les espces viennent
disparatre. Apprhen-
der lesmcanismesqui prsident
leur naissanceest plus dlicat. Une
rcente tude, mene sur deux
espces de poissons du lac Apoyo,
auNicaragua, dmontresans qui-
voque lexistence dun processus
de spciation cest--dire de
sparation dune espce en deux
espces lles qui ntait, jusqu
prsent, que timidement suspect
par la thorie de lvolution.
La doxa veut quau sein dune po-
pulationhomogne deuxgroupes
doivent trespars doncsoumis
des pressions environnemen-
tales diffrentes , et ce, pendant
de longues priodes, pour quils
se diffrencient en deux espces
distinctes. Cest la spciation dite
allopatrique .
Rcemment publis dans la revue
Nature, les travaux de lquipe
dAxel Meyer, chercheur audpar-
tement de biologie de luniversit
de Constance (Allemagne), per-
mettent de dmontrer que deux
espces peuvent provenir dune
mme population sans avoir t
gographiquement spares. Ce
que les spcialistes appelleront un
cas de spciationsympatrique .
Axel Meyer et ses coauteurs ont
jet leur dvolusur unpetit lac de
cratre(5kmdediamtreet 200m
de profondeur) dont ils savent
quil sest form voil moins de
vingt-trois milleans. Dans celac se
trouvent deuxespces decichlids
Amphilophus zaliosus et Amphi-
lophus citrinellus dont lanalyse
gntique montre quelles sont
monophyltiques, cest--dire is-
sues duneseuleespce. Les cher-
cheurs montrent galement quil
y a des diffrences cologiques
entreles deuxpopulations, grce
lanalyse des contenus stomacaux
et des dents pharyngiennes , ex-
plique Herv Le Guyader, profes-
seur de biologie de lvolution
luniversit Paris-VI.
Pourquoi deux espces se sont-
elles diffrencies ? Lanctre
commun aux deux espces est
un poisson de rivire qui vit
de faibles profondeurs, explique
M. Le Guyader. Ces animaux arri-
vent dans un lac dont la colonne
deau est de 200 mtres environ.
Il va donc y avoir des possibilits
de spcialisation : certains vont
demeurer prs des rives, dautres
vont exploiter les niches colo-
giques situes en profondeur.
Ainsi, alors quaucune barrire
physique ne les spare, les deux
groupes vont voluer trs rapi-
dement en moins de vingt-trois
mille ans vers deux espces
distinctes.
Slection disruptive
Les auteurs de ces travaux invo-
quent un processus de slection
disruptive . Soumis un change-
ment denvironnement, un petit
groupe peut voluer de faon
favoriser les individus situs aux
deux extrmes dun trait de ca-
ractre par exemple les gants
et les nains.
Le processus de spciation sym-
patrique, dmontr dans le rgne
animal sur un unique cas, est-il
une banalit de lvolution ? Il
pourrait expliquer, par exemple,
la grande diversit despces de
poissons des rcifs coralliens. Ou
encore lexistence de plusieurs
centaines despces monophyl-
tiques de cichlids dans certains
grands lacs africains, comme
par exemple le lac Tanganyika.
Ces travauxouvrent la porte la
recherche dautres cas de spcia-
tion sympatrique, pour pouvoir
ventuellement constater la gn-
ralit de ce phnomne, estime
Michel Veuille, directeur dudpar-
tement systmatique et volution
du Musum national dhistoire
naturelle(MNHN). Cest important
pour connatre lhistoire des es-
pces, cest galement important
dans le cadre des proccupations
actuelles sur la conservation de la
biodiversit.
Stphane Foucart
(23 mai 2006)
POURQUOI
CET ARTICLE ?
Lexemple des poissons du lac
ApoyoauNicaraguadmontre
que deux espces peuvent
natre dune mme popula-
tion sans aucune sparation
par une barrire physique.
Ce processus particulier de
spciation est li dans ce cas
lexistencedediffrentesniches
cologiques auseindunmme
milieu. Ltude de ces espces
met envidencedes recherches
actuelles pour valider certains
processus volutifs envisags
thoriquement.
Lvolution lenvers
L
arbregnalogiqueduvivant
sestconstruitparlaformation
continuedenouvellesespces.
Laspciationdmarrelorsquedeux
populations dune espce sisolent
en sadaptant progressivement
des environnements distincts. Si
au cours du temps ces populations
sediffrencient aupoint dedevenir
incapables deproduireunedescen-
dance viable et fertile, elles consti-
tuent alors deuxespces spares.
En accumulant des mutations,
alatoires, les espces se transfor-
ment et sadaptent leur environ-
nement. Celles qui ne sadaptent
pas voient leur dmographie d-
cliner, puis disparaissent, comme
le fameux dodo de lle Maurice. Il
existe cependant une autre faon
de disparatre, moins visible et
moins intuitive : la dspciation.
Lorsqueles ressources cologiques
samenuisent, deuxespces surs,
rcemment spares dans des
niches cologiques diffrentes,
peuvent tre contraintes cohabi-
ter et fusionner pour ne plus for-
mer quune seule espce, hybride
des deuxpremires.
Ce processus de spciation len-
vers, mconnu, vient defairelobjet
dunetudepublieenfvrier dans
la revue Nature. Des chercheurs
suisses, emmens par Ole Seehau-
sen, se sont intresss auxespces
dugenre Coregonus, cousines des
saumons, qui peuplent les nom-
breux lacs alpins forms aprs la
dernire glaciation, il y a douze
mille ans. Ces diffrentes espces
de poissons occupent des niches
cologiques bien diffrencies
dans ces lacs : les parties littorales
peu profondes pour les unes, o
elles se nourrissent de crustacs et
pondent en hiver ; les zones plus
profondes pour les autres, qui vi-
vent aux dpens du zooplancton
et pondent t comme hiver.
Au cours du XX
e
sicle, les activi-
ts humaines ont profondment
altr lcologie des lacs. Les rsi-
dus dengrais (azote et phosphore)
qui y ruissellent ont entran la
prolifrationdalgues. Lorsquelles
meurent, ces algues se dposent
au fond des lacs et augmentent
la croissance des bactries, ce qui
appauvrit les eaux profondes en
oxygne. Ce phnomne, leutro-
phisation, force les Coregonus des
eauxprofondes remonter vers la
surface, cohabiter avec dautres
espces et donc partager leurs
sites de ponte, favorisant la forma-
tiondhybrides.
En comparant les gnes et les
formes de milliers de poissons
collects rgulirement depuis
plus de quatre-vingts ans dans ces
lacs, les chercheurs rvlent quau
gr de la pollution croissante
plusieurs espces de Coregonus
ont disparupar dspciation. Des
espces autrefois distinctes sont
aujourdhui fusionnes. Parado-
xalement, ladmographie globale
des poissons est inchange, ce
qui superciellement a pulaisser
croire la bonne sant des lacs. En
ralit, en quelques dcennies, la
diversit des espces sest effon-
dre dans les lacs pollus.
Onpense souvent que les change-
ments volutifs noprent que sur
de trs grandes chelles de temps,
au minimum des milliers dan-
nes. On voit ici pourtant de
quelle manire, en quelques di-
zaines dannes, un groupe des-
pces de poissons a pu tre bou-
levers par des changements
cologiques brutaux et profonds.
La dspciation est un phno-
mne vraisemblablement sous-
estim dans la mise en danger de
la biodiversit. La protectionde la
diversit ncessite de prserver
les habitats mais galement de
comprendre les rouages de lvo-
lution des espces.
Nicolas Gompel,
BenjaminPrudhomme
(12 mai 2012)
POURQUOI
CET ARTICLE ?
Cet article trs rcent envisage
lun des processus de dispari-
tion des espces par hybrida-
tion de deux espces proches
qui se retrouvent partager
le mme milieu. Les auteurs
parlent de dspciation .
Cependant, ce terme tant ab-
sent desprogrammesofciels,
il fautlutiliseravecprcaution.
Larticlemetlaccentsurladif-
frenceentreladmographie
(nombre dindividus) et la di-
versit des espces (nombre
despces diffrentes), et met
en relation les disparitions
despces avec les activits
humaines polluantes.
Gntique et volution Gntique et volution
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
24
LESSENTIEL DU COURS
25
LESSENTIEL DU COURS
Un regard sur
lvolution de lhomme
La place de lhomme parmi
les primates
Les primates partagent des caractres communs,
dont un pouce opposable aux autres doigts de
la main, des ongles plats et des orbites en faade
(favorisant une visionbinoculaire). Les grands singes
dits hominodes se caractrisent par labsence de
queue et la pratique aumoins partielle de la bipdie.
Except lhomme, qui a colonis toute la plante, les
primates vivent dans les rgions intertropicales du
globe (Australie exclue).
Ltude des caractres anatomiques ne permet pas
forcment de prciser les liens de parent au sein
dugroupe des hominodes dont fait partie lhomme.
Les comparaisons chromosomiques et molculaires
sur de nombreux gnes montrent que les plus
proches parents de lhomme sont les chimpanzs.
Elles ont permis dtablir larbre phylogntique
des primates et destimer les dates de divergence
entre les lignes.
Les plus lointains anctres communs lensemble
des primates seraient apparus il y a 65 50 millions
dannes (Ma) en Afrique. Les hominodes seraient
apparus il y a 23 millions dannes, les hominids
(dont font partie les gorilles, les chimpanzs et les
hommes), il y a 10 millions dannes. Les primates
partageant lanctre commun le plus rcent avec
lhomme actuel sont les chimpanzs (Pan troglo-
dytes) et les bonobos (Pan paniscus).
Les premiers reprsentants de la ligne humaine,
qui ont donc unanctre communavec lhomme plus
rcent quavec les chimpanzs, seraient apparus il y
a 5 10 millions dannes.
Les caractres distinctifs de lhomme
Lhommesedistingueduchimpanzpar unensemble
de caractres dont certains sont lis la bipdie per-
manente : untrouoccipital avanc, des membres an-
trieurs rduits par rapport auxmembres postrieurs,
unbassincourt et large et des fmurs convergents en
sont des exemples. Par rapport
celui du chimpanz, le crne de
lhommetmoignedunvolume
crbral plusimportant, etsaface
est aplatie. Dautres diffrences
sobservent dans la mchoire et
la dentition. Les tmoignages
dactivitsculturelles(outils, gra-
vures, peintures rupestres, etc.)
sont trs dvelopps. Lorsquun
fossile prsente au moins lun
de ces caractres distinctifs, il
est susceptible dtre plac au
sein de la ligne humaine, sans
L
espce humaine appartient lordre des primates, qui re-
groupe environ 230 espces dune grande diversit et dont
les comparaisons anatomiques et molculaires permettent
de prciser les liens de parent. Le squenage des gnomes de
lhomme et du chimpanz a rvl une identit de plus de 99 %.
Ainsi, les tudes gntiques et lobservation du dveloppement
de lhomme et de son plus proche parent permettent de com-
prendre comment se construisent leurs phnotypes diffrents
partir dun patrimoine gntique trs proche. Lhistoire de la li-
gne humaine, dont lhomme actuel est lunique reprsentant,
montre une volution buissonnante. Sa phylognie reste discute
et rvisable tout moment, au gr des dcouvertes de fossiles.
Primates
Hominodes
Hominods
Hominids
Gorillins (Gorille)
Panins (Chimpanz)
Homme (Homo sapiens)
Australopithecus sp.
Homo sp.
Hominins
autres Primates
Hylobatods (Gibbon, etc.)
Pongids (Orang-outan)
Arbre phylogntique des primates
pour autant tre considr comme un anctre direct
de lhomme actuel.
La construction du phnotype humain
Lanalyse de 97 gnes fonctionnels de lhomme et
du chimpanz a montr une concidence de 99,4 %.
Ltude du dveloppement pr et post-natal chez les
deux espces permet de comprendre comment de si
petites diffrences gntiques peuvent aboutir des
phnotypesdistincts. Danslesdeuxcas, lephnotype
sacquiert sousleffet delinteractionentrelexpression
de linformationgntique et lenvironnement.
Des mutations affectant lexpression des gnes ho-
motiques peuvent induire des ralentissements ou
acclrations de certaines phases du dveloppement,
ayant pour consquence des modications anato-
miques (taille, forme et position dlments du sque-
lette). Ainsi, lorsdudveloppement embryonnairechez
lhomme, lallongement des phases de mise enplace
du systme nerveux conduit un dveloppement
plus important du cerveau que chez le chimpanz.
Lhomme conserve lge adulte des caract-
ristiques qui nexistent que chez les jeunes pri-
mates : on parle de notnie. En particulier le
trou occipital reste central et permet la bipdie
chez lhomme, tandis quil recule au cours du dve-
loppement post-natal du chimpanz quadrupde.
La relation aux autres individus, les interactions
sociales (apprentissages, transmissiondes cultures,
etc.) contribuent faonner ce phnotype dtermin
gntiquement.
Une volution buissonnante
De nos jours, laligne humaine est reprsente par la
seule espce Homo sapiens, mais elle a t prcde
par de nombreuses espces disparues, notamment
des genres Australopithecus et Homo.
Orrorin tugenensis et Ardipithecus kadabba ( 6 Ma)
en Afrique orientale et Sahelanthropus tchadensis
Touma ( 7 Ma) en Afrique centrale sont les plus
anciens hominins dcouverts.
Les australopithques ont vcude 4,2 Ma 1 Ma ;
on trouve de nombreux fossiles en Afrique de lEst.
Les traces de pas dusite de Laetoli enTanzanie attes-
tent de leur bipdie, il y a 3,6 Ma.
Le genre Homo regroupe des espces bipdes avec
unebotecrnienneplus volumineuse, unerduction
de la face et des canines rduites. Homohabilis enest
le premier reprsentant connu, les fossiles les plus
anciens retrouvs en Afrique sont dats de 2,3 Ma.
Homo erectus ( 1,8 0,6 Ma) est le premier
domestiquer le feuet produire et utiliser des outils
plus sophistiqus : les bifaces. Certains groupes
dHomoerectus saventurent hors dAfriqueet partent
la conqute de lEurasie en plusieurs vagues, de
1,7 Ma120000ans. Leur arrive enEurope serait
date de 650000ans.
Homosapiens serait apparuenAfriqueil yamoins de
200000ans et aurait remplacles populations dHo-
mo erectus au cours de ses migrations successives.
Ces groupes produisaient des outils perfectionns
et varis, et enterraient leurs morts. De nombreux
tmoignages artistiques ont t dcouverts.
Homo neanderthalensis ( 80 000 35 000 ans),
retrouvexclusivement enEurope, prsenteuncrne
volumineux et un bourrelet sus-orbitaire pais. Il
fabriquait des outils de pierre taille et enterrait
ses morts. Les raisons de son extinction ne sont pas
totalement lucides. Il a certainement t en com-
ptitionavec Homosapiens, avec lequel des changes
gntiques ont pu avoir lieu.
La ligne humaine montre une volution qui nest
pas linaire mais buissonnante : plusieurs lignes
ont volu et cohabit, voire ont t interfcondes.
Lhistoire rcente montre une rduction de la diver-
sit : Homo sapiens est le seul reprsentant actuel
de ce buisson touffu. Cette rduction de la diversit
concerne plus gnralement les hominodes, dont
certains sont envoiedextinction. linverse, dautres
primates comme les cercopithcodes montrent une
grande diversication lheure actuelle.
Une grande partie des connaissances qui permettent
dtablir les arbres phylogntiques des primates sont
fondes sur des dcouvertes palontologiques. Ces
connaissances sont donc rvisables.
colonne vertbrale
position du trou occipital
rapport volume crnien / face
bassin
longueur relative des
membres et position
de la jambe
Caractres qui distinguent homme et chimpanz
Deux exemples doutils dcouverts proximit de fossiles
de la ligne humaine
ZOOM SUR
La place de lhomme dans le
monde vivant
Lhomme fait partie, avec les
grands singes, du groupe des ho-
minids, qui appartient lordre
des primates. Comme tous les
mammifres, il est unvertbram-
niote et ttrapode, caractristique
partage avec les amphibiens, les
reptiles et les oiseaux. Enn, il par-
tage avec tous les autres animaux
des caractristiques encore plus
anciennes, commelanaturedeses
cellules, qui sont eucaryotes.
NOTIONS CLS
ARBRE PHYLOGNTIQUE
Traduit des liens de parent entre
des espces. Pour chaquecaractre
on dnit deux tats, lun tant
drivdelautre. Lepassagedeltat
ancestral ltat driv dun carac-
tre est une innovation volutive
qui a ncessairement t trans-
mise de gnration en gnration
jusquauxorganismesobservs. Les
nuds de larbre indiquent le der-
nier anctre commun des espces
qui partagent descaractresdrivs
exclusifs. Larbre phylogntique
dit qui partage quoi avec qui et
doncqui est leplusprocheparent
dequi et nonpasqui descendde
qui (G. Lecointre, La construc-
tionde phylognies , APBG, 1995 ;
1 : 109-36).
LES FOSSILES NE SONT
PAS DES ANCTRES
Les fossiles correspondent des
espces relles ayant vcu durant
une priode gologique donne et
ne concident pas avec les nuds
des arbres phylogntiques.
PARENT
Chercher laparentcest chercher
le groupe frre, et non lanctre. Il
sagit, pour uneespcedonne, du-
groupeaveclequel ellepartageune
innovation volutive exclusive :
elle ne la partage avec aucunautre
groupe. Lanctre au sens propre
restera toujours inconnaissable,
mais on peut dduire certains des
caractres quil devait possder, les
caractresdrivsqui dnissent le
groupe (G. Lecointre, op. cit.).
MOTS CLS
HTROCHRONIE
Modication de la dure ou de la
vitesse des phases du dvelop-
pement au cours de lvolution.
HORLOGE MOLCULAIRE
Estimation de la date de diver-
gence entre diffrentes lignes
depuis leur dernier anctre com-
mun par comparaisons molcu-
laires. Pour un gne donn, plus
les squences de deux espces
sont diffrentes, plus la diver-
gence entre les lignes de ces deux
espces est ancienne.
NOTIONS CLS
VOLUTION
BUISSONNANTE
Les nombreux fossiles dcou-
verts ne doivent pas tre consi-
drs comme une succession
despces descendant les unes
des autres. Ces fossiles peuvent
tre des reprsentants de lignes
disparues. Larbre phylogn-
tique des hominins ressemble
un buisson avec de multiples
branches.
HOMO SAPIENS
Sur le plan de lvolution, Homo
sapiens est une espce comme
les autres. Il nest pas labou-
tissement de lvolution, et
ltablissement de ses liens de
parent avec les fossiles de la
ligne humaine doit tre men
avec la mme rigueur que pour
nimporte quelle espce. Ce-
pendant, plus les fossiles sont
rcents et nombreux, plus des
liens de parent prcis peuvent
tre tablis.
ZOOM SUR
Lucy
Cette australopithque dcou-
verte en thiopie en 1974 a 3 mil-
lions dannes. Son squelette a
t reconstitu 40 % grce aux
52 fragments osseux dcouverts.
Les australopithques sont de
petite taille, prsentent une
faible capacit crnienne, sont
bipdes, mais leur dmarche
devait tre chaloupe .
Gntique et volution Gntique et volution
DEUXARTICLESDUMONDECONSULTER
Origines de lhomme
Une histoire rinventer p. 27-28
(Christiane Galus, 30aot 2006)
Neandertal en nous p. 28-29
(Herv Morin, 8 mai 2010)
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
LESSENTIEL DU COURS
26
UN SUJET PAS PAS
27
LES ARTICLES DU
27
Lintitul du sujet
partir des seules informations recueillies par lex-
ploitation du document :
placez sur larbre phylogntique les innovations
volutivesloriginedescaractresdrivsdutableau;
citez les caractristiques du plus rcent anctre
commun lhomme, au chimpanz et au gorille ;
placez lorang-outan sur larbre phylogntique et
prcisez le degr de parent entre lorang-outan et
chacune des autres espces de larbre.
Le document
Lanalyse du sujet
Pourtraitercesujet, il fautavoircomprislaconstruction
dunarbrephylogntiquepartir deltudedescarac-
tres drivs prsents par diffrentes espces.
Proposition de corrig
Les innovations se placent sur les entre-nuds
Labsence de queue est un caractre driv prsent
chez les quatre espces, donc chez leur dernier anctre
commun. Linnovation volutive est placer avant le
nudcorrespondant.
Lafusionprnataledesosdupoignetetlaprsencedun
sinusfrontal sontdeuxcaractresdrivsexclusivement
communs lhomme, au chimpanz et au gorille. Ces
innovationssont placeravant lenudreprsentant le
dernier anctre communde ces trois espces.
Labipdieest uncaractredrivuniquement prsent
chezlhomme: cecaractrenexistaitpaschezledernier
anctre communde lhomme et duchimpanz, il est
placer aprs le nud.
Caractristiques du plus rcent anctre commun
lhomme, au chimpanz et au gorille
Cet anctre communprsentait les caractres drivs
communs aux trois espces : absence de queue,
prsence dun sinus frontal et fusion prnatale des
os du poignet. Il ne prsentait pas le caractre driv
propre lhomme : la bipdie.
Degr de parent entre lorang-outan et les autres
espces de larbre
Lorang-outan partage deux caractres drivs avec
lhomme, le chimpanz et le gorille. Il ne prsente
pas de fusion prnatale des os du poignet. Sa branche
sintercale donc entre le dernier anctre commun de
lensemble des espces et le dernier anctre commun
de lensemble homme-chimpanz-gorille.
Lorang-outan possde avec lhomme, le chimpanz et
legorilledeuxcaractresdrivsetunanctrecommun
quil ne partage pas avec le gibbon (un seul caractre
drivencommun). Il estdoncplusprochedelhomme,
duchimpanz et dugorille que dugibbon.
Partie 2.1 : Liens de parent
au sein des primates
Arbre phylogntique de quelques
espces de primates actuels
complter
Gibbon Homme Chimpanz Gorille
Arbrephylogntiquedequelquesespcesdeprimatesactuels
Orang-
Outan
1
2
3
4
1- Absence de queue
2- Prsence d'un sinus frontal
3- Fusion prnatale des os
du poignet
4- Bipdie
Espces
Caractres drivs
Gibbon Homme Chimpanz Gorille Orang-outan
Absence de queue + + + + +
Fusion prnatale
des os du poignet
+ + +
Prsence
d'un sinus frontal
+ + + +
Bipdie permanente +
Le signe + signie que le caractre driv est prsent, le
signe signie quil est absent.
Ce quil ne faut pas faire
Utiliser uniquement vos connaissances pour
rpondre aux questions sans faire
de raisonnement logique partir du document.
AUTRES SUJETS POSSIBLES AU BAC
SUR CE THME
Partie 2.1 : Positionner des espces de primates
fossiles dans un arbre phylogntique partir de
ltude de leurs caractres.
Partie 2.2 : Discuter de la remise enquestiondun
arbre phylogntique de laligne humaine suite
la dcouverte dunfossile.
Partie 2.2 : Mettre en relation la comparaison de
squencesdecertainsgneset lesdiffrencesmor-
phologiques entre le chimpanz et lhomme.
ZOOM SUR
LA CONTROVERSE IDA
En mai 2009, New York, Darwi-
nius masillae, unfossile de 47 mil-
lions dannes baptis Ida ,
tait prsent comme unpossible
anctre commun lensemble des
singes anthropodes et qualifi
de chanon manquant par ses
dcouvreurs. Les tudes qui ont
suivi cette dcouverte mdiatise
et controverse ont montr quIda
tait en fait un reprsentant dun
groupe anctre des lmuriens.
Les comparaisons de la dentition,
des mchoires et de 360 carac-
tres chez plus dune centaine
de primates a permis de rfuter
l'hypothse initiale. Les caractres
ressemblants chez Darwinius ma-
sillae et les anthropodes seraient
apparus indpendamment par
convergence volutive, sous la
pression du milieu identique.
LE TROUBLE JET PAR LA
DCOUVERTE DE TOUMA
La dcouverte dun fossile peut
poser davantage de problmes
quil nen rsout. Ce fut le cas de
Touma, dcouvert au Tchad en
2001. Dat de 7 millions dannes,
proche de la divergence entre la
ligne humaine et celle des chim-
panzs, Sahelanthropus tchadensis
a sem le trouble enraisondulieu
de sa dcouverte, et de sa mor-
phologie. Bien plus g que les
australopithques, il prsente des
caractres plus humains : une
face peu prognathe, des canines
rduites et une probable bipdie,
alors que sa capacit crnienne et
satailledevaient trecomparables
celles dun chimpanz.
LVOLUTIONNAPASDEBUT
Pour Teilhard de Chardin (1881-
1955), lhominisation est len-
sembledes processus volutifs par
lesquels les hommes ont acquis les
caractres qui les distinguent des
autres primates . Lhominisation
concerne notamment lacquisi-
tion dune bipdie de plus enplus
parfaiteetdunencphaledeplusen
plusvolumineux. Attentiontoute-
foisauxformulationsqui suggrent
une nalit lvolution, lhomme
nenest pas laboutissement.
Gntique et volution Gntique et volution
Origines de lhomme
Une histoire rinventer
Ces dix dernires annes, la paloanthropologie a considrablement progress, grce
aux dcouvertes de nouveaux fossiles et la gntique. Plus elle se dvoile, plus lhis-
toire de lhomme devient complexe.
L
a paloanthropologie, qui
clbre cette anne cent
cinquante ans dexistence,
est en rvolution. Ces dix der-
nires annes ont vu une accu-
mulation de dcouvertes qui
bouleversent les thories, cham-
boulent les modles dmer-
gence de lhomme moderne,
bousculent les ides reues sur
la prtendue unicit de notre
espce. Ltude des nouveaux
fossiles en Afrique de lOuest,
en Europe, en Asie mais aussi
les progrs de la gntique ne
dressent pas le portrait dune
humanit quittant lanimalit
pour progresser linairement
vers Homo sapiens, mais celui
dune grande diversit despces
humaines ayant coexist.
Le chemin parcouru, depuis un
sicle et demi, est considrable.
Cest le Franais Boucher de
Perthes qui, auXIX
e
sicle, fonde
la prhistoire en prsentant
lAcadmie des sciences un m-
moire sur les silextaills dcou-
verts en 1844 dans les terrasses
de la Somme, prs dAbbeville.
Il estime que ces silex, trouvs
ct de restes de mammouths
et de rhinocros, sont de main
dhomme. Son mmoire est
refus, mais il persvre et est
suivi par dautres scientiques
franais et trangers.
Enaot 1856, uncoup de pioche
dans une carrire de la valle
de Neander en Allemagne, prs
de Dsseldorf, fait apparatre
dtranges ossements humains.
Cette dcouverte, suivie par
dautres, met en vidence, pour
la premire fois, lexistence
dune autre humanit, celle
dHomo neanderthalensis. Cest,
stricto sensu, la naissance de
la paloanthropologie. Les
caractristiques particulires
de lhomme de Neandertal
dplairont fortement aux pa-
lontologues. Le Franais Mar-
cellin Boule en fera une brute
image qui colle encore la
peau de notre cousin. Quand,
plus tard, on trouvera les restes
anciens dHomo sapiens notre
espce , bienplus prsentables,
beaucoup formuleront, dune
certaine manire, leur soulage-
ment.
On remonte, aujourdhui, plus
loin. Les plus anciens reprsen-
tants de lespce humaine ont
t dcouverts en Afrique, tels
Homo habilis (2 millions dan-
nes) et Homo ergaster (1,9 mil-
lion dannes). Dautres ont t
mis au jour en Asie et en Gor-
gie (Homo georgicus, 1,8 million
dannes). Actuellement, la plu-
part des paloanthropologues
saccordent penser quHomo
ergaster, grand et taill pour la
course, a t lorigine de la pre-
miremigrationdelhumanit
partir delAfrique. Toutefois, cer-
tains spcialistes, comme Robin
Dennell et Wil Roebroeks, sup-
putent que les premiers grands
explorateurs ont putre les aus-
tralopithques, des pr-humains
qui ont vcu en Afrique entre
4,2 et 2,5 millions dannes. Ces
hominidsmarchaient debout et
possdaient des mchoires puis-
santes munies dedents robustes.
La grande interrogationactuelle
concerne lanctre commun
hominid-chimpanz. On en
ignore tout, faute de fossiles,
et on ne connat pas la date de
la sparation entre les deux
espces. Les paloanthropolo-
gues saccordent pour la xer
vers 7 millions dannes. Cela
permet dintgrer dans la ligne
humaine des tres trs anciens
tels Ardipit hecus kadabba (5,2
5,8millions dannes), dcouvert
en thiopie par Yohannes Hai-
l-Slassi ; Orrorin tugenensis
(6 millions dannes), mis au
jour auKenya par Brigitte Senut
et Thomas Pickford, et Sahe-
lanthropus tchadensis (Touma,
7 millions dannes), rvl par
les quipes de Michel Brunet et
considr, malgr des contesta-
tions, comme appartenant au
rameau humain.
Mais la mi-2006, une tude
gntique, ralise par Nick
Patterson et David Reich du
Massachusetts Institute of Tech-
nology (tats-Unis) et publie
dans la revue Nature, jette un
pav dans la mare en reculant
la date de la spciation entre
hominid et chimpanz entre
6,3 et 5,4 millions dannes. De
plus, estiment les chercheurs,
elle ne sest pas faite enune fois :
aprs stre spares, les deux
espces se seraient retrouves
et hybrides, avant de se sparer
dnitivement.
Lhypothse formule par les
scientiques amricains quils
qualient eux-mmes de pro-
vocatrice partir de ltude de
20 millions de paires apparte-
nant augnome de lhomme, du
chimpanz, dugorille, delorang-
outanet dumacaquepourrait re-
mettre en cause lappartenance
des plus vieux pr-humains
au groupe des hominids. Ce
que contestent bien videm-
ment les paloanthropologues,
et notamment Michel Brunet
(universit de Poitiers, CNRS).
Ce dernier estime quil sagit de
jeuxintellectuels , tandis que
Jean-Jacques Jaeger, professeur
depalontologieluniversitde
Poitiers, estime que lhypothse
nest pas dmontreet laqualie
de go-posie .
En revanche, il est clair, selon
lui, qu on ne sait pas quoi
ressemblaient ces tres .
chaque fois quon dcouvre un
fossile nouveau trs ancien,
poursuit Jean-Jacques Jaeger, on
met en vidence des mosaques
de caractres quon ne connat
pas. Par exemple, Touma, Or-
rorinet Ardipit hecus forment un
stade volutif nouveau incluant
la bipdie. Mais dans quel cadre
celle-ci est-elle apparue ? On
prend aujourdhui ce caractre
commeunesignaturedelhomi-
nisation. Mais cest peut-tre un
attribut ancestral de quelques
singes dAfrique, que certains
dentreeuxont perdu, explique
le scientique.
Comme le souligne un autre
chercheur, Marc Godinot, pa-
lontologue et spcialiste de
lvolution des primates au
Musumnational dhistoire na-
turelle, une bipdie posturale
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
28
LES ARTICLES DU
29 29
POURQUOI
CET ARTICLE ?
Cet article retrace lhisto-
rique des dcouvertes en pa-
loanthropologie, ltude des
fossiles de la ligne humaine
depuis cent cinquante ans et
la dcouverte de lhomme de
Neandertal. Il fait le point sur
lescontroverseset permet ainsi
de comprendre lvolution des
idesetlecaractrervisabledes
thories en fonction des nou-
velles dcouvertes. Dun mo-
dle dvolution linaire, les
recherches ont abouti une
vision trs buissonnante
de lhistoire de lhomme. De
plus, la divergence entre les
lignes des hommes et des
chimpanzs se rvle de plus
en plus rcente mais aussi
complexe : elle divise les
spcialistes. Il faut tre ca-
pable, partir de documents,
dinterprter diffrentes d-
couvertes en respectant les
rgles de ltablissement des
liens de parent, comme pour
nimporte quelle espce, bien
quil sagisse de lhomme.
dans les arbres a pu prcder
une bipdie de marche, an que
le bassin puisse sadapter . Car
il fallait bien que les premiers
hominids aient la capacit de
biense dplacer debout avant de
saventurer sur lesol delasavane.
On ne pourra pas avancer sur
les plus anciens reprsentants
de la ligne humaine tant quon
naura pas des fossiles dats de
10 et 5 millions dannes, soit
avant et aprs le dernier an-
ctre commun, avance Pascal
Picq, matre de confrences au
Collge de France. Cest le seul
moyen de connatre lvolution
de diffrents caractres. La s-
parationentre les espces a sans
doute t plus rcente et plus
complexe quon ne le pense.
Aupassage, Pascal Picqgratigne
les paloanthropologues, aux-
quels il reproche davoir une
visiongradualiste de lvolution
humaine. Mmesi ontrouvedes
fossiles de 8 9 millions dan-
nes, je suis sr quonles mettra
dans la ligne humaine et non
dans celle des grands singes.
Le plus ancienanctre commun
hominid-chimpanz nest pas
le seul sujet de dbat actuelle-
ment. On sinterroge toujours
sur les causes de la disparition
de lhomme de Neandertal, une
espce admirablement adapte
son environnement glaciaire.
Son dclin inexorable, il y a
trente mille ans, a-t-il t pro-
voqu par la concurrence avec
Homosapiens, par la maladie ou
par tout autre chose ?
Une troisime interrogation
concernelepetit hommedcou-
vert, en 2003, sur lle de Flors
enIndonsie. Vieux de dix-huit
mille ans et dot dune stature
de 1 m, il possdait une capacit
crbrale de 380 400cm
3
, plus
proche de celle du chimpanz
oude laustralopithque que de
celle de Lucy (3,2 millions dan-
nes). En raison de ses caract-
ristiques, ses dcouvreurs lont
class dans une nouvelle espce
humaine : Homo oresiensis. Et
certains paloanthropologues
pensent que ce petit tre, issu
dun Homo erectus rest isol
pendant longtemps dans lle,
a vu sa taille diminuer, comme
cela est arriv pour de nom-
breuses espces animales.
Mais cette explication est
conteste par plusieurs tudes.
La dernire en date, publie le
21 aot dans les Proceedings of
the National Academy of
Sciences (PNAS) et ralise par
une quipe de chercheurs indo-
nsiens, australiens et amri-
cains, estime que lhomme de
Flors serait en ralit issu
dHomo sapiens pygmes, at-
teints de plusieurs anomalies,
dont la microcphalie.
Christiane Galus
(30aot 2006)
Neandertal en nous
Gntique. LADN dHomo neanderthalensis a parl : il se serait appari Homo sapiens
venu dAfrique, avant de disparatre il y a 30 000 ans. Le gnome des populations
dEurasie porte la trace de ce mtissage.
I
l y a du Neandertal en nous.
Du moins si nous sommes
non africains . Dans ce
cas, 1 % 4 %de notre matriel
gntique a pour origine Homo
neanderthalensis. Nous nous
croyions simples cousins, issus
dun anctre commun. Nous
nous dcouvrons aussi mtisss
avec cet humaindisparu. Cest la
conclusionlaplus spectaculaire
tire de ltude de lADNprlev
sur trois os de nandertaliens
vieux denviron 40 000 ans,
issus dune grotte croate.
Pour lapremirefois, legnome
nuclaire dun homme fossile
est squenc, hauteur de
60 %. Son analyse est publie,
au terme de quatre annes
defforts, dans la revue Science,
vendredi 7mai, sous ladirection
de Svante Pbo, de lInstitut
Max-Planck danthropologie
volutionniste de Leipzig. Cest
lui que lon doit la premire
analyse gntique dun nan-
dertalien, en 1997. Il sagissait
dADNmitochondrial (ADNmt),
transmis par la mre, dextrac-
tion bien plus aise que lADN
nuclaire. En 2004, M. Pbo et
ses collgues avaient concluque
cet ADNmt ne rvlait aucun
croisement entreHomosapiens,
lhomme moderne et celui de
Neandertal.
Conclusion aujourdhui invali-
de par les mmes chercheurs,
qui necachent dailleurspasleur
surprise. Lquipe tait contre
cette hypothse, ce qui, dune
certaine manire, renforce nos
rsultats , rappelleDavidReich
(MIT et Harvard), cosignataire
de ltude. Et de rappeler les
prcautions mthodologiques
prises pour se prmunir contre
les contaminations par lADN
des exprimentateurs, limi-
ner celui des microbes prsents
dans les chantillons, qui repr-
sentaient audpart plus de95 %
des lments squencs.
Il a aussi fallu trouver les
moyens de comparer lADN
attribu Neandertal celui
dhommes actuels un Fran-
ais, un Han chinois, un Papou
de Nouvelle-Guine, un San
dAfrique de lOuest et un Yo-
ruba dAfrique de lOuest et
celui dun chimpanz. Pour-
quoi ? Parce que les variations
gntiques non partages avec
Neandertal clairent lhis-
toire volutive de lhomme ,
rpond Ed Green, premier si-
gnataire de larticle de Science.
Ces comparaisons ont donc
mis en vidence la prsence de
1 % 4 %dADNdorigine nan-
dertalienne dans le gnome de
nos contemporains dEurasie,
alors quun tel hritage est ab-
sent chez les deux Africains.
Impossible ce stade de dcrire
le rle fonctionnel de cet ADN
transfuge. Mais sa mise en vi-
dencemet nunecontroverse
scientique vieille de plusieurs
dcennies sur dventuels croi-
sements entre les deuxespces.
Mais sans doute faut-il rappeler
qui est Neandertal, et pourquoi
il constitueuncousinsulfureux.
En 1856 sont dcouverts prs
de Dsseldorf une calotte cr-
nienne et des os longs bientt
attribus (1864) une nouvelle
espce, baptise Homo nean-
derthalensis. En 1859, la publi-
cation de LOrigine des espces
par Darwin banalise lhomme
auseindurgneanimal. Double
choc pour les contemporains. Le
mythe de la Gense est branl,
la cration compte soudain
une nouvelle espce humaine.
Pire, ondcouvre, LaChapelle-
aux-Saints (Corrze), en 1908,
un squelette de Neandertal
enterr par ses congnres : ces
primitifs avaient-ils le souci de
lau-del ?
Depuis lors, ces questions nont
cess de tarauder les paloanth-
ropologues, et dautres se sont
poses au l des dcouvertes :
Neandertal est le seul homi-
nid apparu en Europe, il y a
environ 400 000 ans, descen-
dant dHomo plus archaques
venus dAfrique. A-t-il t ray
de la carte, il ya 30000ans, par
Homo sapiens, lui-mme sorti
dAfrique il y a 50 000 ans ?
Quid de la nature des relations
entre les deux espces, qui ont
pu avoir des contacts pendant
quelques millnaires ?
Ces questionnements sac-
compagnent de querelles sur
le degr d intelligence et
dhumanit quil convient dac-
corder ce perdant de lvolu-
tion. Avec ses bourrelets sus-
orbitaireet sonchignoncrnien,
lincroyable robustesse de son
squelette et de sa musculature,
mais songros cerveau, ntait-il
quun balourd simiesque ?
Cest aussi une crature poli-
tique : certains chercheurs en
font un bon sauvage, victime
des murs colonialistes incoer-
cibles de lhomme moderne,
dautres soulignent les proximi-
ts entre les deux espces, qui
se seraient fondues lune dans
lautre. Cest un sujet explo-
sif, car la notion de diffrence
biologique est dlicate manier
ds lors quon sapproche de
lhumain, notelepaloanthro-
pologueJean-JacquesHublin, lui
aussi delInstitut Max-Planckde
Leipzig, pour qui la recherche,
fconde, de diffrences avec ces
espces disparues na rien de
commun avec celle dune hi-
rarchie entre dhypothtiques
races au sein de lespce
humaine.
La gntique vient donc dap-
porter son grain de sel dans
ces polmiques souvent vives.
Doit-on continuer parler de
deux espces, si leur hybrida-
tion a donn lieu une descen-
dance aussi fertile ? Je laisse
dautres le choix de se querel-
ler ce sujet , rpond Svante
Pbo, qui juge la discussion
assez vaine et se contente de
souligner quil y a bien eu croi-
sements, mme une chelle
modeste. Les populations en
jeunedpassaient pas quelques
milliers dindividus. Il a suf
de quelques dizaines daccou-
plements pour que lempreinte
de Neandertal marque notre
patrimoine gntique.
Certains chercheurs nont pas
attendu les gnticiens pour
se forger une conviction. Cest
le cas du paloanthropologue
Erik Trinkaus (Washington
University, Saint-Louis, Mis-
souri), pour qui il existe dj
en abondance des preuves pa-
lontologiques montrant des
ux de gnes entre nander-
taliens et hommes modernes,
rsultant de labsorption des
populations des premiers par
lexpansion des seconds, il y a
environ 40000ans .
Sauf, rpond Jean-Jacques Hu-
blin, que la gntique suggre
des mtissages plus anciens,
puisque tous les Eurasiens, et
pas seulement les Europens de
lOuest, en portent la marque :
Probablement ont-ils eu lieu
du ct de lactuelle Isral, o
la frontire entre populations
de nandertaliens et dhommes
modernes atuctuanteentre
120 000 et 50 000 ans , soit
avant quHomo sapiens nen-
tame sa marche triomphale
travers le globe.
Ce qui a fait ce succs volutif,
Jean-Jacques Hublin espre le
trouver non seulement du ct
des os saspcialit oudelar-
chologie, mais aussi de celui
des gnes. Larticle de Science
ouvre en effet dautres pers-
pectives fascinantes : disposer
dugnome de Neandertal offre
un aperu de ce qui fait la sp-
cicit dHomo sapiens.
Lquipe internationale a ainsi
mis envidencequelques gnes
uniques notre espce, qui ont
la particularit de jouer un rle
dans le dveloppement cognitif
et le mtabolisme nergtique.
Lorsquils sont endommags,
certains sont impliqus au-
jourdhui dans la trisomie 21,
lautisme ou encore la schi-
zophrnie. Un autre lest dans
laformeducrne, delaclavicule
et de la cage thoracique.
Erik Trinkaus est critique vis-
-vis de cette expdition de
pche aux gnes, qui ne
nous disent rien sur la biolo-
gie ou le comportement de
Neandertal. Bruno Maureille
(CNRS-UMR Pacea, Bordeaux)
juge au contraire trs int-
ressante cette nouvelle tape
de la palogntique, qui va
sintresser aux produits des
gnes, les protines. Il note ce-
pendant que les moyens tech-
niques voluent beaucoup
plus vite que les capacits de la
communaut scientique en
intgrer les rsultats .
La palognticienne Eva-
Maria Geigl (Institut Jacques-
Monod) souligne, elle aussi,
quil faudra du temps avant
que tout le monde soit convain-
cu. Dautant quelagnomique
nen est qu ses dbuts : On
va certainement sapercevoir
que la diversit gntique est
beaucoup plus importante
quon le pense , dit-elle, tant
entreles espces humaines qu
lintrieur de celles-ci.
Leipzig, on en est dj la
prochaine tape pour caract-
riser les gnes qui font le
propre de lhomme : le squen-
age du gnome dHomo sa-
piens contemporains des der-
niers Neandertal est lanc.
Herv Morin
(8 mai 2010)
POURQUOI
CET ARTICLE ?
Les tudes gntiques sur
les hommes de Neandertal
fossiles rvlent des mtis-
sages avec Homo sapiens,
qui taient encore dmentis
par les recherches menes en
2004. Larticle retrace lvo-
lutiondes ides sur ce cou-
sin sulfureux et montre
queles rcentes dcouvertes
remettent enquestionbeau-
coup de certitudes sur lhis-
toire de la ligne humaine
encrant lasurprisedessp-
cialistes eux-mmes. Malgr
le scepticisme de certains
scientiques, les recherches
actuelles portent sur les sp-
cicits gntiques qui font
le propre de lhomme.
Gntique et volution Gntique et volution
LES ARTICLES DU
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
30
LESSENTIEL DU COURS
Gntique et volution 31
LESSENTIEL DU COURS
Gntique et volution
Les relations entre organisation
et mode de vie, rsultat de
lvolution : lexemple de la vie
fixe chez les plantes
Les changes entre la plante
et son milieu
La plante ralise des changes dnergie et de ma-
tire avec son milieu de vie travers des surfaces
dchanges externes trs dveloppes, en lien avec
son mode de vie x. La partie souterraine de la
plante est constitue de lappareil racinaire, qui
assure lancrage duvgtal dans le sol et labsorption
de leau et des ions minraux du sol pour permettre
la photosynthse. Les racines des plantes prsentent
des structures spcialises dans labsorption: lespoils
absorbants. Unpoil absorbant est form dune cellule
trs allonge de lpiderme racinaire. Lensemble des
poils absorbants de la plante constituent une surface
dchange importante entre la plante et le sol.
Les feuilles, portes par les tiges, sont majori-
tairement formes de cellules chlorophylliennes,
contenant des chloroplastes. Ces organites captent
la lumire et synthtisent, partir du dioxyde de
carbone et de leau, des glucides lors de la photo-
synthse. Les feuilles constituent alors une vaste
surface dchange avec latmosphre. Les changes
de molcules gazeuses de CO2
, dO2 et dH2O entre
les feuilles et latmosphre seffectuent au niveau
des stomates, dont lorice lostiole peut tre ou-
vert ou ferm. Les feuilles sont galement respon-
sables de la capture de lnergie lumineuse. Ainsi
lorganisation fonctionnelle des plantes prsente
dimportantes surfaces dchanges en relation avec
leur mode de vie x. De plus la croissance continue
des plantes participe augmenter les possibilits
dchanges entre la plante et son milieu.
La circulation de matire
au sein de la plante
Dans la plante, des changes de matire
la diffrence des animaux qui sont gnralement mo-
biles dans leur environnement, ce qui leur permet de
chercher leur nourriture, de se protger des agressions de
lenvironnement ou dchapper leurs prdateurs , la plante
est constitue de racines ancres dans le sol portant des tiges
feuilles qui se dveloppent en milieu arien. Comment lorga-
nisation des plantes eurs est-elle adapte leur mode de vie
x linterface entre le sol et latmosphre ?
Fleur
Lumire(UV)
Feuille
Collet
Racinesecondaire
Racine
Bourgeon
H
2
O
H
2
O
Ionsminraux
O
2
CO
2
Tige
Lorganisation dune plante eur et les changes avec son
milieu
sont indispensables, dune part, entre les
parties souterraines, lieu de labsorption
des ions minraux et de leau, et, dautre
part, entre les parties ariennes, lieudex-
positionlalumireet dchanges des mo-
lcules gazeuses. Ces changes dematires
dans la plante ont lieudans deuxrseaux
distincts de vaisseauxconducteurs. Lexy-
lme transporte la sve brute, compose
deauet dions minraux, des racines vers
le reste de laplante. Lephlometransporte
lasvelabore, richeenglucides synthti-
ssparlaplante, desorganeschlorophylliens
producteurs, dont lesfeuilles, verslesautres
organes consommateurs de la plante (les racines mais
aussi les organes comme les fruits, les tubercules, etc.).
Les mcanismes de dfense des
plantes
Les plantes prsentent une grande varit de dfense
contre les agressions du milieu. Elles sont capables
dersister auxvariations saisonnires. Par exemple,
les arbres des rgions tempres rsistent aufroidde
lhiver enperdant leurs feuilles et enprotgeant leurs
bourgeons sous des cailles protectrices. Les plantes
annuelles passent la saison froide sous forme de
graines qui assurent la reprise de la vgtation aux
beauxjours. Chez les plantes adaptes auclimat m-
diterranen, laprsencedepoils et decuticulepaisse
auniveaudes feuilles limite les risques de dshydra-
tation en t. Les plantes prsentent galement des
mcanismes dedfense contreleurs prdateurs : des
piquants (comme les cactus), des feuilles pineuses
(comme le chardon). Certaines plantes synthtisent
des toxines les rendant impropres la consomma-
tion. Ainsi, la consommation, par les antilopes, de
feuilles dacacias entrane une augmentation de la
productiondetanins auniveaudes feuilles, qui limite
le broutage de ces herbivores. Enn, certaines plantes
sont mme capables dmettre des signaux attirant
des insectes parasites de leurs prdateurs. Cest le cas
de la vesce des champs, qui produit dunectar attirant
des fourmis, lesquelles attaquent les acariens et les
insectes herbivores qui sont ses prdateurs.
La reproduction des plantes eur
Les eurs sont constitues de diffrentes pices
orales, situes sur des cercles concentriques, appels
verticilles. De lextrieur vers lintrieur de la eur
se trouvent les spales, les ptales, les tamines
et le pistil. La mise en place des verticilles est sous
le contrle de gnes du dveloppement oral. Les
tamines et le pistil constituent les organes repro-
ducteurs de la plante. Le pistil, renfermant les ovules
ou gamtes femelles, forme lorgane reproducteur
femelle.
Les tamines, qui abritent les grains de pollenconte-
nant les gamtes mles, forment lorgane reproduc-
teur mle.
La plupart des fcondations chez les plantes sont
croises : elles ont lieuentredeuxgamtes provenant
de deux individus diffrents de la mme espce.
Dans ce cas, le mode de vie x des plantes impose
un transport du pollen appel pollinisation. La
pollinisationpeut tre effectue par le vent, leauou
des animaux, majoritairement des insectes. Dans le
cas dune collaborationentre unanimal pollinisateur
et la eur, des adaptations souvent trs troites entre
ces derniers sont observes. Par sa couleur, sa forme,
la scrtion de nectar ou encore son parfum, la eur
attirespciquement lanimal capabledelapolliniser.
Lanimal pollinisateur prsente des adaptations mor-
phologiques laccrochagedupollen. Ces adaptations
sont le rsultat dune covolution entre linsecte
pollinisateur et la plante eur.
Aprs la fcondation des ovules contenus dans le
pistil par les gamtes mles des grains de pollen, les
ovules fconds se transforment en graines, tandis
que la eur se transforme en fruit.
De par le mode de vie xe de la plante, les graines
sont responsables de la dispersion de lespce et de
lacolonisationdenouveauxmilieux. Les graines ou
les fruits contenant les graines sont dissmins par
levent, leauouencore les animauxqui les transpor-
tent accrochs leur corps ou en les consommant et
en les rejetant dans le milieu par leurs excrments.
Laussi, il peut yavoir unecollaborationtroiteentre
lanimal dissminateur et la plante, suite une co-
volution.
Coupe longitudinale de tige montrant les
vaisseaux conducteurs
Lorganisation de la eur : A: coupe transversale dune eur ; B : vue
de dessus ; C : diagramme oral
MOTS CLS
DISSMINATION
Transport des graines ou des
fruits contenant des graines
permettant le dveloppement
dune nouvelle plante partir
dune graine. Les principaux
agents dissminateurs sont le
vent, leau et les animaux. La dis-
sminationassure le maintiende
laire de rpartition de la plante
mais galement son extension.
PLANTE
Vgtal terrestre dont les racines
et les tiges feuilles sont relies
par des tissus conducteurs (le
xylme et le phlome transpor-
tant respectivement la sve brute
et la sve labore). Parmi les
plantes, ondistingue notamment
les plantes eurs, ou Angios-
permes, et les plantes sans eurs,
contenant enparticulier les Pino-
phytes (pins, sapins).
POLLINISATION
Transport des grains de pollen
des tamines jusquau pistil
dune eur. Les principaux vec-
teurs de transport du pollen sont
le vent, leau et les animaux.
STOMATE
Structure situe au niveau de
lpiderme des feuilles, princi-
palement sur la face infrieure
de ces dernires, qui permet
les changes de gaz (CO2
, O2 et
H2O) entre les feuilles et latmos-
phre. Un stomate est constitu
de deux cellules pidermiques
spcialises, les cellules stoma-
tiques, encadrant un orifice,
lostiole. Louverture de lostiole
permet le passage des gaz entre
latmosphre et la cavit situe
sous lostiole, la chambre sous-
stomatique. La chambre sto-
matique est le lieu dchanges
gazeux entre lair et les cellules
chlorophylliennes environ-
nantes, comme les cellules du
parenchyme lacuneux. Les
plantes ont la capacit de rguler
louverture de leurs stomates en
fonction des conditions clima-
tiques, ce qui leur permet de
contrler la perte deauauniveau
des feuilles.
ZOOM SUR...
LE SEXE DES PLANTES
La majorit des plantes portent
des fleurs hermaphrodites ou
bisexues, contenant la fois un
organe reproducteur mle, les
tamines, et un organe reproduc-
teur femelle, le pistil. Mais chez
certaines espces dites mo-
noques , comme le mas, un in-
dividuportedes eurs unisexues,
cest--dire des eurs mles (avec
des tamines mais sans pistil) et
des eurs femelles (avec un pis-
til mais sans tamines) sur un
mmepied. Enn, chez les espces
dioques, comme le saule, chaque
individu ne porte quun type de
eurs unisexues, soit mles soit
femelles.
AUTOFCONDATION ET
FCONDATION CROISE
Chez certaines espces comme le
bl, il peut yavoir autofcondation.
Lautofcondation, impossible
chez les espces dioques, a lieu
entre ungrainde pollenet lovule,
issus dun mme individu. Cette
autofcondation peut entraner
une homozygotie (prsence de
deux allles identiques pour un
gne donn) importante chez
les descendants et rduire ainsi
les possibilits dadaptation des
individusauxvariationsdescondi-
tions de lenvironnement. Chez de
nombreuses espces monoques
ouhermaphrodites, lautofconda-
tionest rendue impossible par des
barrires morphologiques (chez
les orchides, une structure ana-
tomique, le rostre, spare le pistil
des tamines), temporelle (les
tamines et le pistil natteignent
pas leur maturitenmmetemps,
comme chez le mas, o le pollen
est mis alors quelepistil nest pas
encore rceptif) ou, le plus sou-
vent, gntique (des mcanismes
dincompatibilit gntique ren-
dant impossiblelautofcondation
chez de nombreuses espces).
La fcondation croise, qui a lieu
entre deuxeurs situes sur deux
plantes diffrentes dune mme
espce, est donc courante chez
les plantes et prsente lavantage
dtre lorigine dune diversit
gntique.
TROISARTICLESDUMONDE
CONSULTER
Le nombre et la varit des insectes
pollinisateurs diminuent en Europe de
manire importante p. 34
(Christiane Galus, 23 juillet 2006)
Les pesticides font perdre le nord aux
abeilles p. 35
(MartineValo, 31 mars 2012)
Des jachres transformes en garde-
manger pour les abeilles p. 35-36
(Jean-PierreTenoux, 28.12.10)
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
LESSENTIEL DU COURS
Gntique et volution 32
UN SUJET PAS PAS
33 Gntique et volution 33
Lintitul complet du sujet
Les yuccas sont des plantes originaires du continent
amricain. Dans leurs pays dorigine, les pieds portent
de nombreux fruits charnus, gnralement comes-
tibles. Un horticulteur install en rgion parisienne a
leprojet decultiver desyuccasdestinslaproduction
de fruits qui seront commercialiss sous forme de
prparations culinaires et cosmtiques. Dans un pre-
mier temps, il prvoit lamiseenculturedenombreux
individus, andeslectionner, par descroisements, les
pieds produisant les fruits avec les qualits requises.
Ensuite viendra la productionenmasse des fruits.
On vous demande de poser un regard critique sur le
projet de lhorticulteur.
Montrez que le projet est techniquement ralisable
conditionderespecter certaines contraintes (quevous
prciserez) et deprendreencompteles consquences
ventuelles sur la biodiversit.
Votre rponse sappuiera sur lexploitation du dossier.
Aucunetudeexhaustivedesdocumentsnestattendue.
Les documents
Document 1 : Lareproductionsexue chez les yuccas
Dansleurscontresdorigine, lafcondationdesyuccas
est ralise par des espces de papillons absentes en
Europe. Certaines espces deyuccas ont timportes
avec succs en Europe ds le milieu du XIX
e
sicle.
Elles poussent sans difcult dans les jardins ou en
pots dans la plupart des rgions. Parmi elles, le yucca
glauque est sans doute le plus robuste car il supporte
des gels svres de lordre de -20 C, voire beaucoup
moins. En Europe, les yuccas portent des eurs mais
ne donnent jamais de fruits. Ils se reproduisent uni-
quement par multiplication vgtative.
Document 2 : Inuence de linsecte teigne du yucca
(Tegeticulayuccasella) sur laproductiondefruits par
les yuccas glauques enAlberta(tats-Unis)
Document 3 : Yucca glauque (Yucca glauca) et teigne
duyucca(Tegeticulayuccasella), deuxespcesenvoie
de disparition?
Document 4:
PollinisationduYuccaaloifoliapar linsecte
Pronubayucasella
Dessins extraits de The Popular Science Monthly, Edouard
L. Youmans, 1882
Partie 2.2 : La culture
en Europe de yuccas
originaires dAmrique
Source : rapport du COSEPAC, Canada, 2002
Source : site du ministre de lcologie canadien
La teigne du yucca et le yucca glauque
Nom : teigne du yucca (Tegeticula yuccasella) et yucca
glauque (Yucca glauca)
Description : La teigne du yucca est un petit papillon au
corps brun et la tte blanche, dont lenvergure des ailes
varie de 18 28 mm.
Le yucca glauque est une grande plante vivace qui peut
atteindreunehauteur de100cmet qui produit unegrappe
de eurs blanches son extrmit.
Habitat : Alberta
Situation actuelle : La teigne du yucca est en voie de
disparition (expose une disparition imminente de la
plante ou du pays). Le yucca glauque est une espce
menace(ellepourrait treenvoiededisparitionsi aucune
mesure nest prise pour enrayer les facteurs susceptibles
de la faire disparatre de la plante ou du pays).
Mesures de rtablissement : Ces deux espces sont
interdpendantes, chacune ayant besoin de lautre pour
survivre. La teigne du yucca se nourrit exclusivement
de graine de yucca glauque, qui ne sont produites que
lorsque la plante est pollinise par les teignes adultes.
Femelle de Pronuba yuccasella en train de pondre dans le
pistil de la eur de yucca
Proposition de corrig
Unecontrainte: linterdpendanceentrelesyuccaset
leur insecte pollinisateur
Les yuccas originaires du continent amricain et
cultivs en Europe ne portent pas de fruits, car les
espces dinsectes responsables de leur pollinisation
sont absentes (document 1). Eneffet, le yuccaglauque
est infod une pollinisation effectue par un
insecte particulier, la teigne du yucca. Cette insecte
ne se nourrit que des fruits de cette plante, qui ne
sont produits que lors la pollinisation effectue par
les teignes du yucca adultes (document 3). En effet,
en absence dufs de teigne prsents dans le pistil
des eurs de yucca, cest--dire enabsence de teignes
adultes ayant pondu, les yuccas glauques ne produi-
sent aucunfruit. Enrevanche, enprsence de teignes
de yucca, une quantit optimale dufs pondus dans
le pistil correspond un nombre maximal de fruits
forms (document 2). Une autre espce de yucca, le
Yucca aloifolia, dpend pour sa pollinisation dune
autre espce dinsecte appele Pronubayucasella. Cet
insecte prsente unorgane spcialis lextrmit de
labdomen, lovipositeur, lui permettant de pondre
ses ufs directement dans les ovaires de la eur du
Yucca aloifolia (document 4). Ainsi la covolution a
abouti des spcialisations anatomiques pousses
de linsecte pollinisateur (forme de lovopositeur)
et de la plante (forme de lovaire), do une forte
interdpendance. Comment lhorticulteur peut-il
raliser la pollinisation des yuccas en Europe pour
raliser la fcondation et obtenir des fruits ? Il peut
envisager dintroduire les insectes pollinisateurs
ddis auxespces de yuccas choisies. Il devra veiller
quilibrer les populations de pollinisateurs et de
yuccas pour optimiser son rendement en fruits.
Mais lhorticulteur peut galement envisager un
mode trs diffrent de pollinisation, lui vitant de
faire appel aux insectes pollinisateurs infods
aux yuccas : il peut recourir la pollinisation
manuelle, comme dans la culture de la vanille.
Une autre contrainte : les conditions climatiques
Lhorticulteur doit choisir ses espces de yuccas en
tenant compte des conditions climatiques, pour
russir leur adaptation leur nouvel environne-
ment : par exemple, le yucca glauque est capable
de supporter des gels svres (document 1).
Des contraintes cologiques lies lintroduction
de nouvelles espces
Lintroduction de nouveaux insectes peut prsen-
ter des consquences cologiques : les espces
ainsi introduites peuvent se rvler invasives et
dsquilibrer les cosystmes environnants. Pour
protger les cosystmes existants, lhorticulteur
peut envisager le confinement des espces nou-
velles. Dun autre ct, ces introductions pour-
raient contribuer sauver des espces en danger :
la teigne du yucca glauque est en voie de dispari-
tion et le yucca glauque est menac (document 3).
Dans tous les cas, les solutions techniques rete-
nues et commercialement viables doivent sac-
compagner dune tude rigoureuse de leurs im-
pacts cologiques.
Coupe transversale dans le pistil de la eur de yucca
A: emplacement de lovipositeur ; B : position des ufs
Ce quil ne faut pas faire
Analyser de manire intgrale les documents
dans lordre donn, sans dgager les contraintes
techniques pour lhorticulteur et les
consquences cologiques.
AUTRES SUJETS POSSIBLES AU BAC
SUR CE THME
Partie 2.2 : tudes de documents
Les liens entre la morphologie et lanatomie de la
plante et sonmode de vie xe linterface entre le
sol et latmosphre.
La lutte contre les prdateurs et les variations des
conditions dumilieu.
Les modalits de la pollinisation et de la disper-
siondes graines oudes fruits.
Pices gnitales de Pronuba yuccasella
Extrmit de labdomen prolonge par un ovipositeur, organe
spcialis permettant de pondre les ufs directement dans les
ovaires,aucontactde lovule
ZOOM SUR...
LES SURFACES
DCHANGES
DES VGTAUX
Le mode de vie x de la plante
et la pauvret de son milieu de
vie (eau et ions en faible quan-
tit dans le sol, CO2 peu abondant
dans latmosphre) donnent lieu
au dveloppement de vastes
surfaces dchanges. La surface
moyenne dchange de nutri-
ments, deau et dions minraux
dun animal rapporte la masse
de lanimal est denviron 3 m
2
/kg,
alors quunvgtal dveloppe une
surface dchange deau et dions
rapporte la masse du vgtal
de plusieurs centaines de m
2
/kg.
LES ASSOCIATIONS
SYMBIOTIQUES
DES RACINES
Chez de nombreuses plantes, la
surface dabsorptionracinaire est
fortement augmente grce une
symbiose avec des champignons :
les mycorhizes. Ces champignons
fournissent la plante enlments
de la solution du sol, comme les
phosphates, tandis que la plante
fournit les champignons en
glucides. Chez certaines plantes
comme les Fabaces, une sym-
biose existe avec des bactries
situes au niveau de nodosits
racinaires. Ces bactries du genre
Rhizobiumsont capables de xer
le diazote de lair et de fournir
des composs azots la plante,
qui, en change, la nourrit en
molcules carbones, comme des
glucides.
LES GNES DE
DVELOPPEMENT
FLORAL
Lexistence de plantes mutantes
prsentant des anomalies du d-
veloppement oral montre que
celui-ci est sous le contrle de
gnes de dveloppement. Ainsi,
chez Arabidospsis thaliana, plante
trs tudie en laboratoire, une
mutation dans le gne Agamous
entrane des eurs dont le pistil et
les tamines sont remplacs par
des ptales. Une mutation dans
le gne Apetala est lorigine de
eurs sans ptales.
ZOOM SUR...
LA COVOLUTION
La covolution se dfinit comme
lvolution coordonne de deux
espces en relation troite lune
avec lautre.
Il existe une vritable collabo-
ration entre les animaux polli-
nisateurs et la plante laquelle
ils sont souvent infods. Par
exemple, certains papillons pol-
linisateurs prsentent un organe
de succion extrmement long et
effil, qui sadapte exactement
lperon nectarifre form
par le ptale de lorchide quils
pollinisent. Grce cet organe de
succion, les papillons rcoltent
le nectar situ au fond de lpe-
ron nectarifre et se chargent
en pollen, quils vont emporter
vers dautres fleurs de la mme
espce. Il sagit par exemple de
lorchide Zaluzianskya microsi-
phon, que lon trouve en Afrique
du Sud et qui est pollinise par
la mouche Prosca ganglbaueri.
FRUIT OU LGUME ?
Les haricots verts sont des l-
gumes, mais le concombre, la
tomate, la courgette sont des
fruits !
Dans le langage courant est ap-
pel lgume tout aliment
accompagnant un plat au cours
dun repas. Mais en botanique le
terme lgume dsigne uni-
quement la gousse forme par
le fruit dune famille de plantes
laquelle appartiennent le pois,
les haricots, la lentille, etc., que
lon appelle parfois les lgumi-
neuses . Ainsi, en appliquant
ces dfinitions botaniques, de
nombreux lgumes consom-
ms comme les haricots verts, la
tomate, la courgette sont en fait
des fruits car ils contiennent des
graines.
Dautres lgumes , comme
les pois et les lentilles, sont
en fait des graines. Et de nom-
breuses autres parties vgtales
sont galement utilises pour
lalimentation : des racines (la
carotte), des tiges modifies en
organes de rserve (les pommes
de terre), etc.
abdomen
ovipositeur
ovaire
ovule
A
B
LESSENTIEL DU COURS UN SUJET PAS PAS
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
34
LES ARTICLES DU
35
LES ARTICLES DU
35
Les pesticides font perdre
le nord aux abeilles
P
lus la recherche scienti-
que avance, moins il de-
vient possible de contester
la nocivit des pesticides sur
lenvironnement. Pour son rle
dclaireuse en territoire de plus
en plus min, labeille a dj fait
lobjet de nombreux travaux.
Deux tudes publies dans la re-
vue Science du 29 mars ont opt
pour des conditions ralistes
plutt que pour des expriences
de laboratoire. Elles soulignent
limpact de la famille dinsecti-
cides la plus rpandue dans le
monde : les nonicotinodes, qui
agissent sur le systme nerveux
central des insectes.
Lquipe franaise codirige par
Mickal Henry de lInstitut natio-
nal de la recherche agronomique
(INRA) et Axel Decourtye, colo-
gue pour le rseau des instituts
des lires agricoles et vgtales
(ACTA) a plac avec de la colle
dent uneminusculepucederadio-
identication (RFID) sur le tho-
rax de 653 abeilles mellifres. Les
chercheurs ont voulu vrier si,
comme certains apiculteurs en
avaient formul lhypothse, les
produits contenant du thiam-
thoxampeuvent leur faire perdre
le sens de lorientation.
Le Cruiser notamment, fabriqu
par le groupe suisse Syngenta et
destin au mas et au colza, les
rend-elles incapables de rejoindre
leur ruche ? Aprs avoir observ
les allers-retours de leurs prot-
ges, lquipe a constat que 10 %
31 % de celles qui taient intoxi-
ques, mme faible dose, ne
retrouvaient pas leur chemin. Ce
qui claire en partie le syndrome
deffondrement des colonies. Car,
loin de leur ruche, elles meurent
trois fois plus que le tauxnormal.
Ces rsultats sont contests par
Syngenta. Mais le ministre de
lAgriculture a annonc, jeudi
soir, quil envisageait dinterdire
le Cruiser, expliquant attendre
un avis de lAgence nationale de
scurit sanitaire (Anses) sur
ltude de Science dici n mai,
avant la nouvelle campagne de
semences en juillet .
La deuxime recherche mene en
cosse sintresse au bourdon
sauvage, en dclin lui aussi. Bien
que le Bumbus terrestris tienne un
rle essentiel dans la pollinisation
des fraises, myrtilles et tomates,
lui et ses congnres sauvages
sont bienmoins connus. Penelope
Whitehorn et Dave Goulson, bio-
logistes de luniversit de Stirling,
ont confront des colonies en
dveloppement de faibles doses
de limidaclopride, prsent, entre
autres, dans le Gaucho. Aubout de
sixsemaines, les nids contamins
avaient pris 8 % 12 % moins de
poids que les colonies tmoins, les
bourdons stant moins nourris.
Surtout, les nids avaient produit
85 %moins de reines.
Martine Valo
(31 mars 2012)
POURQUOI
CET ARTICLE ?
partir de 2006, les tudes
scientifiques ont tabli avec
certitude une diminution im-
portante des populations et
de la biodiversit des insectes
pollinisateurs. Aprs ce constat,
il est devenu urgent de com-
prendre lorigine de cette sur-
mortalit an, si cela est pos-
sible, dy remdier. Depuis de
nombreuses annes, les apicul-
teurs ont incrimin lactiondes
pesticides, trs couramment
utiliss enagriculture. Larticle
rapporte les conclusions de
deux tudes scientiques qui
dmontrent la responsabilit
de deuxinsecticides, le Cruiser
et le Gaucho, dans le dclin
des populations dinsectes
pollinisateurs. Lhypothse
qui prvaut actuellement pour
expliquer le dclindes insectes
pollinisateurs est lhypothse
multifactorielle : les insectes
pollinisateurs seraient affaiblis
par unenvironnement qui leur
serait dfavorable (nombreux
produits phytosanitaires,
attaques par des parasites,
etc). Aujourdhui en France, le
Gaucho et le Cruiser sont inter-
dits cause leur toxicit pour
les insectes pollinisateurs. Les
apiculteurs sont dans lespoir
dune amlioration de la sant
de leurs ruches, qui, si elle a
lieu, ncessitera sans aucun
doute du temps.
Gntique et volution Gntique et volution
L
es insectes pollinisateurs
sont indispensables la
reproduction des plantes
eurs donnant des graines,
qui reprsentent 80 % du rgne
vgtal terrestre : volant de
fleur en fleur pour recueillir
le pollen (l'lment fcondant
mle), ils le transportent vers
le stigmate d'une eur femelle,
ce qui permet la fcondation.
Or, depuis quelques annes, les
scientiques pensent que ce ser-
vice gratuit offert par la nature
depuis 140 millions d'annes,
est menac par la baisse de la
biodiversit.
Une enqute mene par Jacobus
Biesmeijer et WilliamKunin (uni-
versit de Leeds, au Royaume-
Uni) et une quipe de chercheurs
britanniques, allemands et ner-
landais conrme, dans la revue
Sciencedu21 juillet, quelamenace
est srieuse. En tudiant diff-
rentes zones en Grande-Bretagne
et aux Pays-Bas, les scientiques
ont constat que les abeilles sau-
vages paient le plus lourd tribut,
avec une baisse de 52 % de leur
diversit dans le premier cas et de
67 %dans le second, par rapport
la situation prcdant les annes
1980. En revanche, la situation
est moins catastrophique pour
les mouches pollinisatrices, dont
le nombre a dclin de 33 % en
Grande-Bretagne mais augment
de 25 %aux Pays-Bas.
Les chercheurs ont aussi regard
quelle tait l'inuence de cette si-
tuationsur les plantes visites par
les insectes. Ils ont ainsi constat
qu'en Grande-Bretagne 75 plantes
sauvages qui ncessitent d'tre
pollinises par des insectes ont vu
leur distributiondiminuer, tandis
que 30 autres, pollinises par le
vent oul'eause sont, aucontraire,
rpandues davantage. Aux Pays-
Bas, seules les plantes pollinises
par les abeilles sauvages ont dcli-
n. Jacobus Biesmeijer et William
Kunin suspectent donc un lien de
cause effet entre le dclindes in-
sectes pollinisateurs et des plantes
pollinises, sans pouvoir prciser
quel est l'lment moteur de cette
situation : l'volution des modes
de culture, l'utilisation des pro-
duits chimiques dans l'agriculture
ou le changement climatique ?
Ils sont inquiets car, quelle que
soit la cause retenue, l'tude sug-
gre fortement que le dclin de
quelques espces peut dclencher
une cascade d'extinctions locales
parmi d'autres espces associes .
Pour Guy Rodet, entomologiste
l'Institut national de la recherche
agricole (INRA) d'Avignon (d-
partement sant des plantes et
environnement), cet article est
important car c'est le premier
mettre scientiquement en vi-
dence le dclin des insectes pol-
linisateurs. Et cela avec un grand
nombre de donnes et sur une
longue chelle de temps. Les
auteurs de l'article de Science ont
en effet travaill sur un million
d'enregistrements raliss dans le
pass par des naturalistes et dont
certains remontent au rgne de
la reine Victoria. En appliquant
des techniques destines rendre
ces donnes comparables entre
elles, les chercheurs ont divis la
Grande-Bretagneet les Pays-Bas en
carrs de10kmdectet compar
la diversit des insectes pollinisa-
teurs avant et aprs 1980. Cette
date a t retenue car il y a eu de
grands changements dans l'agri-
culture pendant cette priode.
Cette tude a t mene dans le
cadre du programme europen
Alarmdestinvaluer les risques
encourus par la biodiversit ter-
restre et aquatique. Une enqute
similaire a t engage en France
par l'quipe de recherche sur la
pollinisationentomophiled'INRA-
Avignon, mene par BernardVais-
sire et qui a pour objectif d'va-
luer ledclindes pollinisateurs sur
une priode de temps trs courte.
Dix parcelles ont t retenues
pour raliser l'tude, rparties en
France, enAllemagne, enPologne,
en Sude, et au Royaume-Uni.
La baisse de la diversit des in-
sectes pollinisateurs peut avoir
divers effets. Elle peut d'abord
se traduire par un changement
du paysage, explique Guy Ro-
det, car il y a un risque de voir
disparatre diffrentes espces
de plantes . Plus grave encore,
nous pouvons avoir des dif-
cults pour produire des fruits
et des lgumes, bienqu'onpuisse
dans certains cas pallier la baisse
des pollinisateurs sauvages par
des insectes issus d'levages,
prcise Guy Rodet. Mais pour
des grandes surfaces cultives,
comme aux Etats-Unis, cela
risque de coter trs cher.
Christiane Galus
(23 juillet 2006)
Le nombre et la varit
des insectes pollinisateurs
diminuent en Europe de
manire importante
POURQUOI
CET ARTICLE ?
Les insectes sont un maillon
essentiel de la reproduction de
nombreuses plantes eurs,
parmi lesquelles des plantes
cultives. On estime ainsi que
35 % du tonnage mondial dali-
ments dorigine vgtale pro-
viennent decultures dpendant
dinsectes pollinisateurs et le
service conomique rendu par
ces insectes pollinisateurs est
valu plus de 150 milliards
deuros par an. Lagriculture et
lalimentation humaine sont
donc largement dpendantes
de ces insectes pollinisateurs.
Dans les annes 1990, dans
plusieurs rgions du monde,
les apiculteurs ont commenc
salarmer dune augmentation
importante de la mortalit de
leurs populations dabeilles.
Larticle, crit en 2006, pr-
sente les premires tudes me-
nes en Europe, qui ont tabli
avec certitude la diminution
dunombreet delabiodiversit
des insectes pollinisateurs. Les
scientiques sont larecherche
de lorigine de cette volutionet
sinquitent des consquences
envisageables : ils observent
dj une modification de la
rpartition des plantes pollini-
ses par les insectes et sinter-
rogent sur limpact possible sur
les plantes cultives.
Des jachres transformes en
garde-manger pour les abeilles
En Franche-Comt, 700 hectares ont t sems de plantes trs nutritives pour aider
les butineuses passer lhiver.
C
ombien dabeilles en
hi bernati on survi -
vront-elles au froid ? En
Franche-Comt, beaucoup
plus quailleurs , espre Ray-
mond Borneck. 86 ans, lan-
cien prsident de la Fdration
mondiale dapiculture (Api-
mondia), retir dans son Jura
dorigine, se bat toujours pour
ses protges . En lien avec
les agriculteurs et les chasseurs,
lhomme y plaide la cause des
jachres apicoles ou cultures
interstitielles exprimentes
localement par le rseau Biodi-
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
versit pour les abeilles. Sur le
plan nancier, linitiative, dun
cot de 180 000 euros sur trois
ans, est soutenue par le conseil
rgional, dont sonls Marc pr-
side le groupe des lus Europe
cologie-Les Verts. Sans lien
de cause effet , se hte de
prciser celui-ci.
Lide de ce projet, labellis par
le ministre de lcologie au
titre de lanne 2010de la biodi-
versit, est simple : fournir aux
colonies de butineuses domes-
tiques, durant larrire-saison,
lalimentationla plus quilibre
et laplus richepossiblepour leur
permettre ensuite de passer lhi-
ver. En Franche-Comt, rgion
pilote, plus de 700 hectares ont
tconvertis enzones dintercul-
tureset semsdespcesconnues
pour leur valeur nutritive.
Les abeilles dhiver ne sont
pas biologiquement sem-
blables celles qui travaillent
lt, souligne M. Borneck. Elles
ne vivent en moyenne quun
mois et demi, contre cinq ou
six mois pour ces dernires.
Leur mettre disposition, par
exemple, certaines varits
de tournesol donnant un pol-
len trs suprieur dautres
permet de leur faire gagner
jusqu un mois de longvit.
Sarrasin, avoine, vesce, phalcie,
sainfoin, lierres, trfle blanc,
de Perse et incarnat, mlilot,
lotier cornicul rendent plus
rsistantes les ruches installes
dans un rayon denviron trois
kilomtres autour des parcelles.
Limportant est de leur offrir
une oraisonplus tale dans le
temps, donc un garde-manger
avec des ressources en nectar et
pollensufsantes pour accrotre
leurs dfenses immunitaires
et la production de miel , r-
sume Pierre Testu, animateur
du rseau Biodiversit pour les
abeilles.
Les agriculteurs qui prtent
leurs sols pendant six mois y
trouvent leur avantage. Entre
deux rcoltes dorge oude mas,
le terrain ne reste pas nu, ces
plantes empchent lapparition
de mauvaises herbes, limitent
lrosion et apportent de la
matire organique , tmoigne
Ins Jacquet, exploitante Ri-
gney (Doubs). Les espces et les
mlanges utiliss pour ces ro-
tations automnales sont au-
toriss par la politique agricole
commune(PAC) sans drogation
sur les surfaces en jachre, car
les espces sont non invasives
et certies.
Le 18novembre, le conseil rgio-
nal de Franche-Comt a ritr
son appui lopration, inscrit
dans une convention plurian-
nuelle relative au dveloppe-
ment des couverts . Sign avec
la Fdration des chasseurs,
qui conduit et coordonne les
actions en faveur de la faune
sauvage et de ses habitats sur le
territoire, ce programme vise
promouvoir la dmarche dans
les milieuxagricoles et apicoles,
accumuler des connaissances
agronomiques sur cettepratique
nouvelle, en dmontrer la fai-
sabilit rglementaire et mesu-
rer lintrt pour les colonies
dabeilles .
Sur la microparcelle de 0,5 hec-
tare qui jouxte le muse de
lAbeille Mesnay (Jura), Ray-
mond Borneck, qui a cr lIns-
titut technique dapiculture de
lINRA, poursuit ses propres
exprimentations. En2011, il ne
mlangera plus les graines. Les
espces seront semes sur des
bandes spares, par types,
pour faire des comptages sur
lintensit des butinages .
Lvolution agronomique des
diffrentes plantes pourra ainsi
tre mieux analyse.
Jean-Pierre Tenoux
(28 dcembre 2010)
36
LES ARTICLES DU
LE DOMAINE CONTINENTAL
ET SA DYNAMIQUE
Noyau
exter ne
Manteau
infr ieur
chellenon
respecte
Manteau
supr ieur
Asthnosphre
Lithosphre
Biosphre&
Hydrosphre
Atmosphre
100-150km
700km
MOHO
GUTENBERG
(2885km)
LEHMANN
(5155km)
Crote ocanique
(6km)
Roches
sdimentaires
Crote continentale
(30-65km)
Noyau
inter ne
Plaquesus-jacente
Magma Volcanisme
500C
0
15
30
60
Profondeur
en km
600C
800C
1000C
1200C
1200C
Plaqueplongeante
Manteau suprieur
L
i
t
h
o
s
p
h
r
e
o
c
a
n
i
q
u
e
SV
SB
E
Crote ocanique
H
2
0
H
2
0
Point A
Manteau
On prcisequelemanteau
est constitudepridotites
et lacroteocanique,
debasaltes et degabbros.
Lgende:
Zonedefusion partielle
delapridotitehydrate
SV Facis schistes verts
SB Facis schistes bleus
E Facis clogites
Fosseocanique
Ocan
Croteocanique
Crotecontinentale
Andsite, rhyolite
L
i
t
h
o
s
p
h
r
e
c
o
n
t
i
n
e
n
t
a
l
e
L
i
t
h
o
s
p
h
r
e
o
c
a
n
i
q
u
e
M
a
n
t
e
a
u
a
s
t
h
n
o
s
p
h
r
i
q
u
e
Plutonde
granodiorite
Manteaulithosphrique
Subduction
S
c
h
iste
s b
le
u
s
c
lo
g
ite
s
Dshydratation
Sismes
Chambremagmatique
formationdumagma
andsitiquepar fusion
partielle
Gntique et volution
POURQUOI
CET ARTICLE ?
Cet article montre comment
plusieurs types dintervenants
se mobilisent en France pour
venir en aide aux populations
dabeilles. Eneffet, durant lhiver,
des abeilles particulires, plus
longue dure de vie que celles
dt, habitent la ruche. Elles se
nourrissent desrservesaccumu-
les dans la ruche avant que le
froidnedeviennetropimportant.
Quand la temprature devient
trop faible (autour de 10 C), les
abeilles ne quittent plus de la
ruche. lintrieur, les abeilles
luttentcontrelefroidenseserrant
les unes contre les autres et en
battant des ailes. Larticle montre
que des apiculteurs, sensibles
la mortalit des abeilles, se sont
mobiliss avec laide des agricul-
teurs pour organiser la culture
de plantes dont le butinage en
automne permet aux abeilles
de prparer efcacement le pas-
sage de la mauvaise saison. Ces
cultures sont ralises sur des
terres qui sont alors en jachre,
cest--dire non cultives. Ce pro-
jetestunbel exempledecoopra-
tionentreplusieurs intervenants
conomiques, scientifiques et
institutionnels.
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
Le domaine continental et sa dynamique Le domaine continental et sa dynamique 38
LESSENTIEL DU COURS
39
LESSENTIEL DU COURS
La caractrisation du domaine
continental : lithosphre
continentale, reliefs
et paisseur crustale
La composition de la crote
continentale
La lithosphre continentale est majoritairement
forme de roches magmatiques (dont le granite et
des roches voisines dugranite), mais aussi de roches
sdimentaires.
Observ lil nu et au microscope polarisant, un
chantillon de granite montre trois types de mi-
nraux : du quartz, des feldspaths (alcalins comme
l'orthose ou plagioclases) et des micas.
Lpaisseur de la crote
continentale
Le forage le plus profond ralis dans la
lithosphre (en pninsule de Kola) est denvi-
ron 12 km et na pas pu traverser la crote. Des
tudes sismiques indirectes permettent destimer
la profondeur de la discontinuit de Mohorovi i
(moho), qui correspond la limite entre la crote
et le manteau. Elle est estime 35 km en moyenne,
mais nest pas identique en tout point du globe.
Au niveau des chanes de montagnes, on observe un
paississement crustal liunraccourcissement dont
les preuves sont visibles sur le terrain.
Les roches compresses, transportes haute al-
titude ou au contraire enfouies sous de grandes
profondeurs, subissent des modications lies aux
variations de pression et de temprature (mta-
morphisme). Les transformations minralogiques
sont accompagnes de changements de texture, les
minraux s'alignant progressivement en feuillets
(schistes, gneiss). Ces conditions peuvent entrqner
une fusion partielle et produire du magma.
Densit et isostasie
Lpaississement crustal observ au niveau des
chanes de montagnes pourrait suggrer unexcs de
masse. Mais en ralit elles exercent une attraction
moindre que celle laquelle on pourrait sattendre.
Ceci peut sexpliquer par la prsence dune racine
crustale, compensant les reliefs en profondeur, en
accord avec les mesures de profondeur du moho.
E
nviron 30 % de la supercie totale du globe terrestre sont
occups par les terres merges. Laltitude moyenne des
continents est de 840 m au-dessus du niveau de la mer,
culminant 8 847 m (mont Everest). Les caractristiques de la
crote continentale permettent dexpliquer son paisseur, son
comportement et ses reliefs.
Composition chimique
Silice
(SiO2)
70%
2,6% 3% 1%
14,5% 8,6%
(Al2O3) (Na2O,K2O)
Alumine Alcalins
Calcium
(CaO) (MgO) (FeO+Fe2O3)
Fer Magnsium
Remarque : il est dusage en gologie
de dsigner les lments par leur
oxyde.
PL
Q
BI
BI
Q
Q
Q
Q
Noyau
exter ne
Manteau
infr ieur
chellenon
respecte
Manteau
supr ieur
Asthnosphre
Lithosphre
Biosphre&
Hydrosphre
Atmosphre
100-150km
700km
MOHO
GUTENBERG
(2885km)
LEHMANN
(5155km)
Crote ocanique
(6km)
Roches
sdimentaires
Crote continentale
(30-65km)
Noyau
inter ne
En effet, la densit moyenne de la crote conti-
nentale est de 2,7, tandis que celle du manteau
est de 3,3.
Il existe en profondeur une surface de compen-
sation sur laquelle les pressions exerces par les
roches sont gales : il y a quilibre isostatique.
Selon le modle dAiry, en fonction du relief
en surface et de la densit des matriaux, des
colonnes de roches sont donc plus ou moins
paisses en profondeur. Au niveau des domaines
continentaux, la racine crustale constitue de
matriaux peu denses stend sur une paisseur
proportionnelle laltitude du massif, la profon-
deur de compensation.
Cet tat dquilibre ralis une profondeur va-
riable de la Terre est lisostasie. Au cours du temps,
lrosion limine progressivement une partie du
massif, ce qui provoque une remonte isostatique
de la racine crustale. Des roches initialement en
profondeur peuvent alors parvenir en surface.
Lge des roches
Alors que la crote ocanique nexcde pas
200 Ma, la crote continentale peut en certains
endroits du globe dpasser 4 Ga. Une roche s-
dimentaire est plus rcente que celle quelle
recouvre (sauf accident tectonique) et peut tre
date en fonction des fossiles quelle contient.
Pour les roches magmatiques, il est ncessaire
de recourir la datation absolue. Cette mthode
de datation est base sur la dcroissance radioac-
tive de certains isotopes radioactifs comme
le
87
Rb. Leur dsintgration en fonction du temps
constitue un chronomtre naturel. Le rubidium
(Rb) et le strontium (Sr) sont prsents en trs
faible quantit (traces) dans les roches magma-
tiques. Ils prsentent diffrents isotopes. Le
spectromtre de masse permet de faire la diff-
rence entre ces isotopes car ils nont pas la mme
masse atomique. Le
87
Rb se dsintgre en
87
Sr,
avec une pri ode de 48, 8. 10
9
ans
( = 1,42. 10
11
an
1
) ; le
86
Sr est un isotope stable.
Lors de la formation dune roche magmatique,
les feldspaths (orthose, plagioclase) et micas in-
corporent des quantits variables de Rb et Sr. Il
nest pas possible de connatre la quantit initiale
disotopes radioactifs au moment de la formation
de la roche. Aprs refroidissement du magma, la
roche forme ne va plus changer avec le milieu
environnant, on parle de fer-
meture du systme. Les iso-
topes voluent spontanment
selon les lois physiques de
dsintgration radioactive : la
quantit dlments pres di-
minue, la quantit dlments
fils augmente. En mesurant les
quantits actuelles des l-
ments pres et fils dans un
chantillon pour au moins
deux minraux diffrents, on
peut tracer une droite (iso-
chrone) et dterminer le
temps coul depuis la ferme-
ture du systme, cest--dire
depuis la cristallisation des
minraux lors de la mise en
place de la roche par refroidis-
sement du magma.
Cascade de lArpenaz
TROIS ARTICLES DU MONDE
CONSULTER
Voyage vers le manteau de la Terre,
dans la crote ocanique
du Pacique p. 41-42
(Pierre Le Hir, 16 avril 2011)
Le voyage sous la Terre du satellite
Goce p. 42
(Christiane Galus, 17 mars 2010)
La structure interne des Pyrnes
dvoile p. 43
(Pierre Rimbert, 7 septembre 1995)
PERSONNAGE
CL
ANDRIJA MOHOROVI I
(1857-1936)
Au dbut du XX
e
sicle, ce
mtorologue et sismologue
croate commena sintresser
la propagation des ondes sis-
miques. Suite au tremblement
de terre de Pokuplje, proximit
de Zagreb, le 8 octobre 1909, il
interprta les sismogrammes et
mit en vidence la discontinuit
entre la crote et le manteau qui
porte son nom (moho).
ZOOM SUR
Les plis et les failles
Les plis se forment par flexion
plastique des roches.
Les mouvements relatifs des
roches le long des failles inverses
tmoignent du raccourcisse-
ment.
Un chevauchement conduit des
ensembles de terrains anciens
en recouvrir de plus rcents.
Un charriage est un chevau-
chement de grande ampleur
pouvant dpasser la centaine
de kilomtres.
Roches
plus rcentes
Roches
plus anciennes
Faille inverse
Force de
compression
Mouvement
des blocs
Pli Faille
Chevauchement Faille
ZOOM SUR...
Le programme ECORS
Le programme tude continen-
tale et ocanique par rexion
et rfraction sismiques (ECORS),
dbut enFrance en1983, consiste
explorer la crote terrestre, ses
variations dpaisseur et sa struc-
ture interne an de dterminer
ses proprits et sonvolution. La
recherche dhydrocarbures, lex-
ploration gothermique et lva-
luation du risque sismique sont
des applications pratiques qui en
dcoulent grce la connaissance
prcise des zones sdimentaires et
des zones fractures.
NOTIONS CLS
LA STRUCTURE INTERNE
DU GLOBE
La Terre est constitue denve-
loppes de compositions chimiques
diffrentes : la crote, le manteau
(constitudepridotites) etlenoyau
(contenant principalement du fer
et du nickel). La discontinuit de
Mohorovi i marquelalimiteentre
la crote et le manteau. La crote
et la partie rigide du manteau su-
prieur constituent la lithosphre,
dcoupe en plaques, reposant sur
lasthnosphre, plus ductile.
LISOSTASIE DAPRS
LE MODLE DAIRY
d1 =densit de la crote continen-
tale = 2,7 ; d2 = densit du man-
teau = 3,3.
Selon le modle dAiry, en fonc-
tion du relief en surface et de la
densit des matriaux, les co-
lonnes de roches sont plus ou
moins paisses enprofondeur. Les
pressions quelles exercent sont
gales au niveau de la surface de
compensation, o il y a quilibre
isostatique.
d1
d2
Schma interprtatif d'une lame mince de granit
observe au microscope. Q : Quartz, BI : Biotite
(mica noir), PL : Feldspaths plagioclases.
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
40
UN SUJET PAS PAS
41
LES ARTICLES DU
Lintitul complet du sujet
Slectionnez la proposition exacte pour chaque
question de 1 6.
1. La crote continentale est principalement
forme :
a) de gabbros et de granites.
b) de roches sdimentaires et de granites.
c) de pridotites.
d) de roches voisines du gabbro.
2. Les diffrences daltitude entre continents et
ocans sont dues au fait que :
a) la crote continentale est moins dense et plus
paisse que la crote ocanique.
b) la crote continentale est plus dense et plus
paisse que la crote ocanique.
c) la crote continentale est moins dense et plus
ne que la crote ocanique.
d) la crote continentale est plus dense et plus ne
que la crote ocanique.
3. Un granitode est une roche :
a) magmatique structure grenue.
b) magmatique structure vitreuse.
c) sdimentaire structure vitreuse.
d) sdimentaire structure grenue.
4. La crote continentale :
a) est en moyenne plus jeune que la crote
ocanique.
b) est plus dense dans les zones de collision.
c) spaissit progressivement dans les zones de
collision anciennes.
d) saltre sous leffet des agents de lrosion.
5. La mthode de datation rubidium-strontium
est fonde sur la dcroissance radioactive du
87
Rb,
qui se dsintgre spontanment en
87
Sr, unisotope
stable. On mesure dans la roche dater les quan-
tits de
87
Rb,
87
Sr et
86
Sr, un isotope stable dont la
quantitest supposeconstanteaucours dutemps.
a) On connat les quantits initiales des lments
pres et ls.
b) Le
86
Sr est un lment ls du
87
Sr.
c) Il est indispensable deffectuer des mesures sur
plusieurs minraux de la roche dater.
d) Cette mthode de datationest adapte auxroches
sdimentaires.
6. Des mesures isotopiques effectues sur des
minraux des granites G1 et G2 ont permis de
construire le graphique ci-dessous. Les droites ont
pour quation:
87
Sr/
86
Sr =( t)
87
Rb/
86
Sr +
87
Sr
0
/
86
Sr.
a) Le granite G1 est plus g que le granite G2.
b) Le granite G2 est plus g que le granite G1.
c) Les granites G1 et G2 sont de mme ge.
d) Le coefcient directeur de la droite isochrone
ne varie pas de la mme faon que t, le temps
coul depuis la fermeture du systme.
Le corrig
1. b), 2. a), 3. a), 4. d), 5. c), 6. b)
Partie 1 : La caractrisation du
domaine continental
Ce quil ne faut pas faire
Se dispenser dune relecture attentive en n
dpreuve car il sagit dun QCM: les rponses
peuvent beaucoup se ressembler et une faute
dinattention est possible.
AUTRES SUJETS POSSIBLES AU BAC SUR CE THME
Partie 2.1
Exploiter des documents pour mettre en relation un modle et des mesures an dexpliquer la notion
disostasie.
Partie 2.1
Identier dans des documents les preuves dun raccourcissement et dun paississement crustal au
niveau dune chane de montagnes.
ZOOM SUR...
LA DATATION ABSOLUE
Ladatationisotopiquearvolution-
n la mesure dutemps engologie.
Encomplment de la datationrela-
tive, qui permet dordonner chro-
nologiquement des vnements
gologiqueslesunsparrapport aux
autres, la datation absolue permet
deleurattribuerunge, enmillions
dannes par exemple.
Cependant lchantillon dater
doit satisfaire certains critres :
il doit sagir dune roche mag-
matique dont on datera la cris-
tallisation par refroidissement
dun magma. En effet, une roche
sdimentaire est notamment
constitue de fragments de reliefs
rods, dont la datation ne don-
nerait pas lge de la formationde
la roche sdimentaire elle-mme
mais de la roche magmatique
dont elle est issue par rosion et
sdimentation ;
lesystmedoit trefermdepuis
la formation de la roche. Aucun
lment ne doit avoir quitt
lchantillon ou y tre entr.
Pour dater un chantillon, il faut
choisir lisotope radioactif le plus
adapt selonsa demi-vie et lge
dterminer. Au-del de dix fois la
demi-vie de lisotope, toute me-
sure est impossible. On mesure
les quantits disotopes radioactifs
prsents dans lchantillonenuti-
lisant un spectromtre de masse.
On procde ensuite aux calculs.
LA DTERMINATION DE
LGE DUNE ROCHE PAR
LA MTHODE
87
RB/
87
SR
87
Rb
87
Sr + e
Soit F la quantit de lisotope
ls prsente actuellement dans
lchantillon, F
0
celle prsente
initialement, P la quantit de
lisotopepre, laconstantecarac-
tristique du couple isotopique :
F = ( t) P + F
0
.
La quantit de
86
Sr tant stable, on
peut utiliser les rapports
87
Rb/
86
Sr
et
87
Sr/
86
Sr.
Ainsi
87
Sr/
86
Sr = ( t)
87
Rb/
86
Sr +
87
Sr
0
/
86
Sr, soit lquation dune
droite de coefcient directeur ( t)
Lecoefcient directeur deladroite
permet decalculer t, letempscou-
l depuis la formationde la roche.
Le domaine continental et sa dynamique Le domaine continental et sa dynamique
Voyage vers le manteau de la Terre,
dans la crote ocanique du Pacique
Au large du Costa Rica, une mission internationale va forer lcorce terrestre sur 2 km
pour prlever des roches profondes, tmoins de lhistoire gologique de notre plante.
mesure que nous
descendions, la
succession des
couches composant le terrain
primitif apparaissait avec plus
de nettet. [] Jamais minra-
logistes ne staient rencon-
trs dans des conditions aussi
merveilleuses pour tudier la
nature sur place. Dans son
Voyage au centre de la Terre,
publi en1864, Jules Verne ima-
gine la jubilationduprofesseur
Lindenbrock et de son neveu
Axel devant ces richesses
enfouies dans les entrailles du
globe .
Unsicle et demi plus tard, len-
thousiasmedeBenot Ildefonse
(Laboratoire de gosciences,
CNRS-universit de Montpel-
lier-II) nest pas moins vif
lide de creuser jusquaux
couches infrieures de lcorce
terrestre, 2 kmde profondeur,
et den remonter la surface,
pour la premire fois, des
roches encore jamais extraites
de leur gangue. Enattendant de
pouvoir unjour traverser cette
pellicule supercielle pour ac-
cder, sous la crote terrestre
paisse de 30kmenmoyenne
sur les continents et de 6 km
sous les ocans , au manteau
qui entoure le noyau de notre
plante et qui reprsente les
deux tiers de sa masse.
Le chercheur est lun des deux
codirecteurs, avec le Britan-
nique Damon Teagle (Centre
national docanographie de
luniversit de Southampton),
de la mission scientique in-
ternationale qui, du 13 avril au
30 juin, va raliser un forage
ocanique dans le Pacique est,
au large du Costa Rica, avec
le bateau amricain Joides
Resolution. Un btiment de
143 m de long, quip dun
derrick de 61 m de hauteur qui
permet de dployer plus de
9 km de tubes de forage dans
les abysses, pour atteindre
lcorce terrestre. bord, un
laboratoire peut effectuer
lanalyse physique, chimique
et magntique des carottes
minrales extraites dusous-sol.
Cette campagne doit pousser
plus avant les forages dj ef-
fectus dans le mme puits,
en 2002 et 2005, jusqu une
profondeur de 1 500 m. Cest-
-dire sans dpasser la crote
ocanique suprieure. Lobjec-
tif est cette fois de descendre
jusqu 2 000 m, au niveau de
la crote ocanique infrieure.
L o les basaltes de surface,
ns du refroidissement rapide
dumagma issude la fusiondes
roches du manteau, cdent la
place aux gabbros, des roches
formes par une cristallisation
plus lente du magma.
Le site prospect se trouve sur
la plaque Cocos, prs dune
dorsale ocanique (zone de di-
vergence entre deux plaques
tectoniques o se produisent
des remontes de magma) o
la formation de la crote, voil
quinze millions dannes, a
t particulirement rapide.
La crote suprieure y est plus
mince quailleurs, ce qui rend
les gabbros plus accessibles.
Lexamen dchantillons
de ces roches permettra de
mieux connatre la structure
de la crote, la faon dont elle
se forme partir du magma
du manteau et leur mode de
refroidissement , explique
Benot Ildefonse.
Ce nest quun dbut. Lexpdi-
tion concide avec le 50
e
anni-
versaire duplus ambitieuxpro-
gramme de forage scientique
de tous les temps : le projet
Mohole, du nom du gologue
croate Andrija Mohorovi i
qui, ds 1909, avait dcouvert
lexistence, la limite entre
lcorce et la partie suprieure
du manteau terrestre, dune
discontinuit se manifestant
par une acclration de la vi-
tesse de propagationdes ondes
sismiques.
Au printemps 1961, au large
de lle mexicaine de Guada-
lupe, les Amricains percrent
plusieurs trous jusqu prs
de 200 m. Mais, faute de tech-
niques de forage adaptes, le
projet fut abandonn cinq ans
plus tard, aprs avoir englouti
plus de 50 millions de dollars
(34,6 millions deuros environ).
Par drision, lentreprise fut re-
baptiseNohole, pas detrou.
Depuis, plusieurs milliers de
forages scientiques ont t
conduits, dans le cadre de
collaborations internationales
dont lactuelle est lIntegrated
OceanDrilling Program(IODP).
Mais aucun na atteint la dis-
continuit de Mohorovi i , plus
simplement appele le moho.
Le plus profondna pas dpass
2 111 m. Lexploitationptrolire
offshore va bien au-del au
large de Cayenne, enGuyane, la
compagnie britannique Tullow
vient dentreprendre unforage
dexploration jusqu 4 300 m
sous le plancher ocanique ,
mais dans des couches sdi-
mentaires recelant des gise-
ments dhydrocarbures, et sans
carottage de roches, qui exige
un quipement plus sophis-
tiqu.
La mission en cours servira
de reprage pour une nou-
velle tentative de creusement
jusquau moho, 4 000 m plus
bas. Elle pourrait tre dcide,
espre Benot Ildefonse, dans la
dcennie venir. Il y faudra des
moyens de forage et de carot-
tage encore plus performants.
Et un nancement internatio-
nal de plusieurs centaines de
POURQUOI
CET ARTICLE ?
Cet article montre que, malgr
les techniques de prospection
indirecte, les tentatives de fo-
rage dans la crote terrestre
des profondeurs de plus
en plus importantes sont
toujours dactualit. Aucun
forage na jamais atteint la
discontinuit de Mohorovi i
(le moho), mais cet objectif
est plus envisageable en mi-
lieu ocanique, o la crote
de 6 km est bien moins paisse
que la crote continentale
(30 km).
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
42
LES ARTICLES DU
43
LES ARTICLES DU
Le domaine continental et sa dynamique Le domaine continental et sa dynamique
La structure interne
des Pyrnes dvoile
Des chercheurs dressent une carte tridimensionnelle de la chane.
I
l y a 65 millions dannes,
lEspagne faisait une en-
tre fracassante au sein
de lEurope. De la collision
entre les plaques tectoniques
Eurasie, au nord, et Ibrie, au
sud, naissait alors la chane
des Pyrnes. Les mcanismes
gologiques ayant conduit
cette formation sont encore
controverss. Annie Souriau,
du Laboratoire de dynamique
terrestre et plantaire, de
Toulouse, et Michel Granet, de
lObservatoire de physique du
globe, de Strasbourg, ont men
une tude sur la structure pro-
fonde des Pyrnes.
Les chercheurs ont utilis pour
cela une technique, trs pro-
metteuse, danalyse du temps
de propagation des ondes
sismiques qui diffre des
systmes (camions vibreurs)
habituellement utiliss par
les ptroliers. Rsultat : une
vritable radiographie dest
en ouest des Pyrnes, inter-
prtable en trois dimensions,
dtaillant avec prcision la
situation des couches golo-
giques jusqu une profondeur
de 200 kilomtres.
Cette tude valide un modle
de formation de la chane
montagneuse, li lhistoire
tectonique mouvemente de
lEspagne. Situe, il ya 200mil-
lions dannes, louest de la
France, alors que la Galice est
en face du Finistre, la plaque
Ibrie entame une lente rota-
tion vers le sud. Tournant au-
tour dunaxe situ louest des
Pyrnes, elle ouvre, sur son
passage, le golfe de Gascogne.
Repousse par la plaque afri-
caine, elle entre encontact avec
lEurope. Lors de cet affronte-
ment tectonique, la plaque Ib-
rie sencastre dans sonvis--vis
europen. La partie suprieure
de la crote slve alors et
forme les Pyrnes.
Ltude des chercheurs a mon-
tr que la partie infrieure de
la crote ibrique a amorc un
plongeonsous sonhomologue
europenne. Elle sest ensuite
enfonce sous lcorce ter-
restre, dans une zone connue
sous le nom de manteau,
jusqu une profondeur de
100 kilomtres. Ces travaux
ont galement mis envidence
la remonte et la perce tra-
vers la crote de deux gros
blocs dorigine profonde. Invi-
sibles ensurface, ils concident
avec des anomalies gravim-
triques connues par ailleurs. La
morsure de ces deux dents
rocheuses travers la couche
sdimentaire pourrait expli-
quer une partie importante
de lactivit sismique de cette
rgion.
Un large rseau
de stations
Pour parvenir ces rsultats,
les chercheurs ont mesur et
analys les temps de propa-
gation des ondes sismiques
provenant de sismes locaux
ou lointains. Chaque couche
traverse par le train dondes
est caractrise par une vitesse
de propagation spcique. Si
le sous-sol est homogne, le
temps de propagation des
ondes ne dpend que de la
distance de leur foyer. En re-
vanche, toute discontinuit de
la structure interne provoque
une avance ou un retard du
front dondes. Ainsi, la traver-
se dune structure vitesse de
propagation leve sera mise
en vidence par une lgre
avance du signal. La compa-
raison des temps darrive
permet de localiser la position
de chaque anomalie.
Mais, pour obtenir une carte
tridimensionnelle grande
chelle dune chane de colli-
sion, il faut disposer dunlarge
rseau de stations capables de
recevoir et denregistrer les
ondes sismiques. La prci-
sion de ltude en dpend. Le
massif pyrnen est quip
dun grand nombre de sismo-
graphes, installs notamment
la suite du tremblement
de terre qui secoua le village
dArette, le 13 aot 1967, au
sud-ouest de Pau (Pyrnes-
Atlantiques). Ce sisme, dune
magnitude de 5,7 sur lchelle
de Richter, dtruisit le petit
village pyrnen et causa la
mort dune personne.
Annie Souriau et Michel Gra-
net ont ainsi bnfici des
donnes issues dune quaran-
taine de stations xes fran-
aises et espagnoles, dont
certaines mises en place dans
des buts trs divers : enregis-
trements sismiques, contrle
des avalanches en Espagne ou
surveillance du gisement de
gaz de Lacq en France. Mais,
regrette Annie Souriau, le
fonctionnement du rseau
pyrnen, gr par lObserva-
toire Midi-Pyrnes, est au-
jourdhui menac dinterrup-
tion. Les stations ncessitent
un entretien rgulier et minu-
tieux. Or, les ingnieurs en
charge de cette tche, bientt
la retraite, ne seront pas
remplacs cette anne.
Pierre Rimbert
(7 septembre 1995)
POURQUOI
CET ARTICLE ?
Cet article dj ancien
dcrit le principe et les
intrts de la prospection
sismique pour la compr-
hensionde la structure pro-
fonde des chanes de mon-
tagnes. Complmentaires de
ltude des afeurements et
des roches, ces techniques
permettent de reconstituer
lhistoire dun relief, selon
des scnarios qui ont volu
depuis 1995 au gr des nou-
velles dcouvertes.
Le voyage sous la Terre
du satellite Goce
En mesurant des variations de gravit, lengin a fourni une carte mondiale dune
frontire gologique, le moho.
L
orsque Isaac Newton
conut la loi de la gravita-
tion universelle, il nima-
ginait pas que, trois sicles plus
tard, des scientiques utilise-
raient les donnes du champ
de gravit terrestre obtenues
par des satellites pour tudier
les profondeurs inaccessibles de
notre globe. Des chercheurs de
lcole polytechnique de Milan
(Italie), dirigs par Daniele Sam-
pietro, viennent eneffet de ra-
liser lapremirecartemondiale
du moho, cette zone frontire
situe entre la crote terrestre
et le manteau suprieur. Ils
ont obtenu ce rsultat en inter-
prtant les donnes recueillies
par le satellite Goce de lAgence
spatiale europenne (ESA), en
orbite 265 kmde la Terre. Cet
engin, lanc en mars 2009, a
fourni des donnes gravim-
triques dune nesse ingale
lchelle de 80 km de len-
semble du globe. Cette image
du moho a t ralise dans
le cadre dun projet (Gemma)
nanc par lESA.
Le moho est le nom de la dis-
continuit dcouverte par
le sismologue croate Andrija
Mohorovi i , alors quil tudiait
la propagation des ondes sis-
miques gnres par le sisme
du 8 octobre 1909 en Croatie. Il
avait dcouvert cette occasion
queles ondes acclraient aprs
le moho. Ce dernier est situ en
moyenne une profondeur de
25 km 60 km sous les conti-
nents et de 5 km 8kmsous les
ocans. Au-del, on trouve le
manteau, puis le noyau liquide
et la graine (le noyau solide),
situe plus de 5 000 km de
profondeur.
Cartographie globale
Toutes ces donnes ont tfour-
nies grceauxondes sismiques.
Car personne contrairement
auxhros deJules Vernenapu
aller sonder lecentredelaTerre.
Les Amricains, les Allemands
et les Russes ont bien tent de
forer le moho, sans rsultat,
bienque les Russes aient atteint
la profondeur de 12 kmen 1989
dans la presqule de Kola.
Jusqu prsent, on possdait
bien des donnes parses sur
les diffrentes profondeurs du
moho, obtenues par les ondes
sismiques ou des donnes gra-
vimtriques locales, dont les
variations retent la diffrence
de densit entre les roches de
la crote et celles du manteau.
Mais lhtrognit de ces in-
formationsdansletempsetdans
lespacesetraduit par degrandes
erreurs dans le rsultat nal ,
estime Daniele Sampietro.
Aujourdhui, grce Goce, les
chercheurs italiens ont ralis
une cartographie globale de
lpaisseur de la crote ter-
restre , explique Michel Dia-
ment, spcialistedegravimtrie
lInstitut dephysiqueduglobe
deParis. Or lacroteterrestreet
le manteausuprieur sont dun
grand intrt pour les scienti-
ques. Cest lendroit ose situe
laplus grandepartiedes proces-
sus gophysiques importants
lis la tectonique des plaques,
tels les sismes, les volcans et la
naissance des montagnes.
Mais comment des donnes
gravimtriques peuvent-elles
fournir des renseignements sur
la structure interne de la Terre ?
Si la Terre tait un oignon,
avec des couches homognes et
sphriques, la pesanteur serait
partout la mme la surface du
globe, prcise Michel Diament.
Or la Terre est htrogne. Il y
a des endroits plus ou moins
denses, si bienque la pesanteur
est variable et rete la rparti-
tion htrogne de la matire
diffrentes profondeurs.
Cette rpartition htrogne
des masses lintrieur de la
Terre contrle galement le
gode, la forme quaurait la
Terre si elle tait recouverte
entirement docans. Et les
scientiques ont dcouvert des
choses surprenantes. En tu-
diant latrajectoiredes satellites,
qui subit linuence du champ
de gravit terrestre, ils ont mis
en vidence une Terre cabos-
se, invisible lil nu, avec
des bosses sur la Papouasie-
Nouvelle-Guine et le Pacique
ouest, et ungrandcreuxassoci
locan Indien. Les six accl-
romtres de Goce ont conrm
ces donnes avec une prcision
ingale.
Christiane Galus
(17 mars 2010)
POURQUOI
CET ARTICLE ?
Cet article rcent montre
lvolution des techniques
de dterminationde la pro-
fondeur du moho, la limite
entre la crote terrestre et
le manteau. Cest unsatellite
mesurant les variations de la
gravit, lies lhtrogni-
t de densit des matriaux,
qui a permis den raliser
une carte mondiale.
millions deuros. Cest ces conditions
que seront mieuxcompris la dynamique
interne de notre plante, la tectonique des
plaques lorigine des sismes tels que
celui du11 mars auJapon, ouencore le rle
de la crote et dumanteauterrestres dans
le cycle du carbone , plaide le chercheur.
Forer jusquaumanteauest le plus grand
d de lhistoire des sciences de la Terre,
dit-il. Paradoxalement, nous en savons
davantage sur dautres plantes, grce aux
missions spatiales, que sur la ntre.
Pierre Le Hir
(16 avril 2011)
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
44
LESSENTIEL DU COURS
Le domaine continental et sa dynamique 45
LESSENTIEL DU COURS
Le domaine continental et sa dynamique
Contexte de la formation
des chanes de montagnes
et disparition des reliefs
Les traces dun ancien ocan
Le mont Chenaillet, situ dans les Alpes, est compos
de roches basaltiques en forme de coussins, qui rap-
pellent les pillow-lavas des dorsales ocaniques. Sous
ce basalte se trouvent successivement des gabbros et
des pridotites : il sagit dune srieophiolitique, ves-
tige dune lithosphre ocanique de 155 Ma, charrie
sur le continent.
Danslazonedauphinoise, prsduBourg-dOisans, des
blocs basculs spars par des failles normales sont
recouverts de formations sdimentaires, rappelant
les dpts des marges passives. Ce sont les vestiges
de la naissance dun ocan.
Lune des causes de la subduction
La lithosphre ocanique est forme de crote oca-
nique (densit 2,9) et de manteau lithosphrique
(densit3,3). Ladensitglobaledelalithosphreoca-
nique dpend des paisseurs relatives de crote et de
manteau. Lalimiteaveclemanteauasthnosphrique
est une limite physique qui dnit le comportement
plus ou moins ductile des pridotites : lisotherme
1 300 C. En sloignant de la dorsale, la lithosphre
ocanique se refroidit, lisotherme 1 300Csenfonce,
et le manteau lithosphrique spaissit : la densit de
lalithosphreocaniqueaugmenteaucoursdutemps.
Entre 30 et 50 Ma, la lithosphre ocanique devient
U
ne partie des terres merges est forme de chanes de
montagnes, anciennes ou toujours en cours de surrec-
tion. Lexemple des Alpes permet dtudier la formation
dune chane de collision dans un contexte de convergence li-
thosphrique. Les traces dun ancien domaine ocanique y t-
moignent de sa fermeture par subduction. Au cours du temps,
les chanes de montagnes, comme tous les reliefs forms la
surface de la Terre, tendent disparatre : la lithosphre conti-
nentale est recycle en permanence.
Ouest Est
LeTaillefer
Coupe des blocs basculs Mure/Taillefer /Rochail
LeRochail
J urassiquesuprieur et crtac(- 96- 154Ma) :
calcaireset marnesammoniteset calpionelles
J urassiqueinfrieur et moyen(- 54- 154Ma) :
calcaireset schistesammonites, blemniteset crinodes
Trias(- 205- 245Ma) : dolomitestrspauvresenfossiles
Ammoniteset blemnites: mollusquesmarinsplagiques(nageant enpleinemer)
Crinodes: organismesbenthiques(fixssur lesfondsmarins)
Calpionelles: organismesunicellulairesmarinsplagiques
Carbonifre(- 295- 355) : schistesnoirset conglomrats
Socleprimaire: rochesmagmatiqueset mtamorphiques
Faille
Vestiges de marges passives dans les Alpes
plus dense que lasthnosphre (densit 3,25) et peut
plonger par subduction, si les conditions y sont favo-
rables. Ceci explique quil nexiste pas de lithosphre
ocanique plus ancienne que 200Ma.
La collision
Lorsquelocansparantdeuxcontinentssestreferm
et quelacroteocaniqueest entirement subduite,
les deux masses continentales saffrontent. Entre
elles subsiste la suture de matriaux ocaniques.
Lessentiel de la lithosphre continentale continue
de subduire, mais la partie suprieure de la crote
spaissit par empilement de nappes dans la zone de
contact entre les deuxplaques.
Des transformations minralogiques
Au cours de la subduction et de lpaississement, les
roches sont soumises des variations importantes
de pression et de temprature, lorigine de la trans-
formationdes minraux: cest le mtamorphisme.
Lesminrauxtrouvsdanslesrochesalpinesmontrent
que celles-ci ont t soumises des pressions et des
tempratureslevesavant derevenir ensurface. Dans
ledomaineinternedesAlpes, lemtamorphismeaune
intensit croissante de louest vers lest (la pression
et la temprature auxquelles ont t soumises les
roches sont de plus en plus leves) : ceci indique
le sens de la subduction. Les basaltes et les gabbros
sont mtamorphiss dans les facis schistes bleus
glaucophane et clogite, tmoignant du mtamor-
phisme haute pression / basse temprature, caract-
ristique de la subduction ayant prcd la collision.
Ces roches mtamorphiques ont unedensitplus le-
ve que les roches d'origine. Ceci a renforc l'enfonce-
ment delalithosphreocaniquedansl'asthnosphre
aucours de la subduction
La disparition des reliefs
Plusieurs orogenses se sont succd au cours de
lhistoire de la Terre : caldonienne (480-420 Ma),
hercynienne (320-260Ma) et alpine. Les reliefs de la
Grande-Bretagne, de la Scandinavie et du Groenland
sont des vestiges de la premire ; les Ardennes, les
Vosges, le Massif central et le Massif armoricain,
de la deuxime. Ces reliefs ont depuis t rods.
Tout relief est donc un systme instable qui tend
disparatre aussitt quil se forme.
Des vnements tectoniques peuvent fragiliser des
dices rocheux. Dautres facteurs, lis au climat et
laltitude, participent lrosion (eau, vent). Les
vgtaux y contribuent galement par la scrtion
de substances acides ou en sinsinuant dans les s-
sures dj formes. Leau est un facteur chimique,
mais aussi physique, drosion : les vagues, les tor-
rents, les glaciers modient les paysages. Charg de
particules de sable, le vent rabote et polit les amas
rocheux. Les blocs et les particules glissent le long
de la pente forme par le relief, puis peuvent tre
transports par leau ou le vent trs loin du lieu
daltration, enfonctionde leur taille. Les sdiments
se dposent lorsquils rencontrent un obstacle ou
un bassin. Ils peuvent alors former une roche sdi-
mentaire.
distancelaxedeladorsale
5km
MOHO
ge
58km
Profondeur
(en km)
isotherme 1300 C
5 M
a
25 M
a
35 M
a
40 M
a
Croteocanique
MOHO
Manteau
lithosphrique
Asthnosphre d=3,25
d=3,3
d=2,9 Hc
Hm
H=
Hcx2,9+Hmx3,3
H
isotherme 1300 C
d=2,9
d=3,3
Dtail de la structure dune lithosphre ocanique /
volution de la lithosphre ocanique au cours du temps
Ophiolite obducte
Eocne
50 Ma
est
Phase de convergence
Crtac sup
80 Ma
Subduction de la lithosphre ocanique
Collision
La naissance dune chane de montagnes
MOTS CLS
DENSIT
Rapport de la masse dun certain
volume dun corps celle du
mme volume deau (ou d'air
pour les gaz).
MARGE
ACTIVE
Limite ocan/continent qui
correspond une frontire de
plaque convergente.
Les marges actives sont le sige
de la subduction et se caractri-
sent par une importante activit
sismique et volcanique.
MARGE
PASSIVE
Zone de transition entre une
crote continentale et une
crote ocanique au sein dune
mme plaque lithosphrique.
Lhistoire de la naissance des
ocans est enregistre au niveau
de leurs marges passives (failles
normales et sdimentation ca-
ractristiques du rifting).
PLAQUE
LITHOSPHRIQUE
Portion de lithosphre dune
centaine de kilomtres dpais-
seur, aux frontires de laquelle
des reliefs (dorsales, chane de
montagnes, fosse ocanique)
et une activit sismique et/ou
volcanique sont observs en
relation avec les mouvements
relatifs des autres plaques.
NOTION CL
La tectonique des plaques
Il sagit dune thorie labore
la fin des annes 1960.
Trois types de mouvements re-
latifs des plaques sont observs
leurs limites :
des mouvements de conver-
gence au niveau des chanes
de montagnes et des zones de
subduction ;
des mouvements de diver-
gence au niveau des dorsales
ocaniques o il y a cration de
lithosphre ocanique ;
des zones de coulissage o il
ny a ni cration ni destruction
de surface lithosphrique.
MOT CL
MTAMORPHISME
Transformations minralogiques
ltat solide dune roche soumise
des variations de pression et de
temprature.
ZOOM SUR...
DES TMOINS
DE LHISTOIRE
ALPINE
Des roches prleves en Vanoise
ont t soumises une pression
de 11 kbars, correspondant une
profondeur de 33 kmet une tem-
prature de 390C.
Des roches prleves au mont
Viso (zone de fort mtamor-
phisme) tmoignent dune
pression de 21 kbars (63 km de
profondeur) et dune temprature
de 550C.
La cosite trouve dans le massif
italien Dora-Maira correspond
du quartz soumis des pressions
suprieures 30 kbars (90 kmde
profondeur).
Onpeut dater les vnements m-
tamorphiqueset lesplacer dansun
diagramme ande tracer untrajet
pression-temprature-temps et de
reconstituer une partie de lhis-
toire de la chane de montagnes.
LROSION
DU GRANITE
Un massif granitique va saltrer,
devenant dugranite pourri et une
arne granitique, un sable dans
lequel des grains de feldspaths,
des paillettes de mica et des grains
de quartz sont identiables. Si le
massif est expos ausoleil, le mica
noir emmagasine la chaleur et se
dilate. Pendant la nuit, la roche
refroidit et se contracte de faon
diffrente dans les parties super-
cielles et profondes. Des ssures
se forment, dans lesquelles leau
depluievapouvoir sinsinuer. Une
successionde gel/dgel fragilise la
roche et largit les ssures.
Boule granitique
Granite pourri
Arne granitique
DEUXARTICLESDUMONDECONSULTER
Des sismes provoqus par le ballet des
continents p. 48
(Christiane Galus, 31 mai 2006)
Le sisme au Japon Pour les sismolo-
gues, ce tremblement de terre nest pas
encore le big one p. 49
(Herv Morin, 12 mars 2011)
Le cycle des roches
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
Lanalyse du sujet
Il sagit de retracer, dans lordre chronologique,
certaines tapes de lhistoire de la formation des
Alpes partir de documents de terrain. Aprs avoir
exploit les indices de lexistence dun ocan alpin
par le pass, la subduction est mise en vidence par
ltude des minraux du mtamorphisme. Enn,
le raccourcissement et lpaississement crustal
permettent de dcrire la collision continentale
ayant succd la fermeture ocanique.
Proposition de corrig
I. Le massif du Chenaillet : des lambeaux dun
ocan disparu ( Exploitation du document 1)
Dans le massif du Chenaillet, nous avons observ
des pridotites mtamorphises ( 2 200 mtres),
puis des gabbros et, partir de 2 400m, des basaltes
en coussins. Il sagit de roches de la lithosphre
ocanique, qui se trouvent normalement au fond
des ocans. Les ophiolites tmoignent de la pr-
sence lemplacement actuel des Alpes dun ocan
aujourdhui disparu. On peut estimer que la mise
en place de cette partie de lithosphre ocanique
date denviron 100 Ma.
II. Les mtagabbros du mont Viso : tmoins
dune subduction passe
(Exploitation des documents 2 et 3)
En Italie, dans le massif du mont Viso, nous avons
trouv des chantillons de mtagabbros, des gab-
bros dune ancienne crote ocanique qui ont subi
du mtamorphisme. Ils contiennent des minraux
du mtamorphisme : grenat, glaucophane et ja-
dite. Ltude du diagramme de stabilit montre
que cette association correspond une tempra-
ture denviron 500 C et une pression de 1 GPa. Il
sagit donc dun mtamorphisme haute pression
basse temprature , caractristique dune zone
de subduction.
Ainsi les mtagabbros du mont Viso tmoignent
dune subduction passe de la lithosphre oca-
nique de locan alpin.
III. Le chevauchement des rochers de Leschaux :
indice dune compression (Exploitation du do-
cument 4)
Enn, nous sommes alls dans le massif des Bornes,
o nous avons observ un contact anormal mon-
trant un chevauchement du crtac infrieur 2 sur
lunit forme du crtac suprieur et du crtac
infrieur 2.
Cette dformation importante des roches rsulte
dune tectonique en compression qui sinscrit dans
le cadre de la collision continentale, lorigine de
la surrection des Alpes.
De plus, nous avons observ dans les calcaires du
crtac infrieur 1 la prsence de fossiles marins de
rudistes (mollusques) et orbitolinids (foramini-
fres), prouvant quils se sont mis en place dans un
domaine ocanique.
46
UN SUJET PAS PAS
47
Lintitul complet du sujet
On cherche montrer comment des donnes de
terrain peuvent permettre de comprendre des v-
nements gologiques anciens.
Vous avez effectu une excursiongologique dans les
Alpes et rapport des chantillons et des photogra-
phies. En vous appuyant sur ces donnes de terrain
et dautres informations prsentes dans le dossier,
rdigez un compte-rendu de votre excursion, illustr
par un ou plusieurs schmas, pour montrer que les
indices recueillis permettent decomprendrecertaines
tapes de lhistoire de la formationde cette chane de
montagnes. Votrerponsesappuierasurlexploitation
du dossier. Aucune tude exhaustive des documents
nest attendue.
Document de rfrence : Carte simplie des Alpes
Les documents
Document 2 : Domaines de stabilit de quelques
associations de minrauxde lacrote ocanique
Document 3 : Observation microscopique dun
mtagabbro dumont Viso (Alpes italiennes) et son
schma interprtatif
Document 4 : ORochers de Leschauxdans les Alpes
(massifs des Bornes, Haute Savoie, Nordde Gre-
noble) et croquis dinterprtation. (Crdit photogra-
phique : ChristianNicollet/ UBPClermont-Ferrand)
Partie 2.2 :
La formation des Alpes
Genve
Grenoble
PROVENCE
UNITS ALPINES :
M
A
S
S
IF
C
E
N
T
R
A
L
JURA
R
h
n
e
Brianon
Massif desBornes
Marseille
Originaires
delamargecontinentaleeuropenne
delocanalpin
sdiments
ophiolites
delamargecontinentaleafricaine
Nice
0 50km
Chenaillet
Mont Viso
Coupe gologique schmatique des ophiolotes du Chenaillet.
LeChenaillet
500m
2600
2400
2200
Altitude (enm)
Pridotitesmtamorphises
Gabbros
Basaltesencoussins
Basaltesenfilons
Brchesmagmatiques
Ce quil ne faut pas faire
Faire une tude exhaustive des documents sans
faire de liens entre les indices recueillis.
Omettre de respecter la forme impose par le
sujet : un compte-rendu dexcursion.
Se dispenser de faire appel ses connaissances.
Ngliger les schmas.
-1
km
CHENAILLET
crote
ocanique
manteau
sdiments
ocaniques
basaltes en coussins
(et basaltes en filons)
pridotites
mtamorphises
gabbros
( et brches magmatiques)
Lophiolite du Chenaillet :
une ancienne lithosphre ocanique
Faille
Chevauchement
Coupe dun chevauchement
Schma du chevauchement indiquant les contraintes
compressives
AUTRES SUJETS POSSIBLES AU BAC SUR CE THME
Partie 2 :
tablir unschma-bilanducycle des matriauxde la crote continentale, avec ousans documents.
Partie 2.1 :
Exploiter des documents et raisonner pour mettre envidence le lienentre lge dune lithosphre ocanique,
sa densit et la subduction.
Partie 2.2 :
Exploiter des donnes cartographiques, des images ou des donnes satellitales pour apprcier lrosion dun
massif actuel.
ZOOM SUR...
LHISTOIRE DES ALPES
Elle commence par la mise en
place dun foss deffondrement
continental, prlude du futur
ocan alpin. Une mer de faible
profondeur sengouffre. Au juras-
sique, locanisation proprement
dite est en marche : locan alpin
spare lEurope de lAfrique. De la
lithosphre ocanique est cre
vers 140Maauniveaudunedor-
sale ; locanest enexpansion. Les
marges de locan sont passives,
des sdiments post-rift tmoi-
gnant de larrt durifting se dpo-
sent. La datation palontologique
de roches contenant des fossiles
marins indique que louverture
ocanique (accrtion) sest drou-
le du jurassique ( 170 Ma) au
crtac suprieur ( 70 Ma), avec
une vitesse dexpansionde lordre
de1 cmpar an.Aucrtacinfrieur,
locan alpin a atteint sa taille
maximale (1 000 km de largeur
estime). Au crtac suprieur, la
fermeture de locan senclenche
par subduction. Aprs disparition
de locan, la marge continentale
europenne est entrane son
tour dans la subduction, puis la
collision proprement dite dbute,
aux alentours de 35 Ma. Un ves-
tigedecroteocaniqueachapp
la subduction, ce sont les ophio-
lites : les nappes ophiolitiques
(Chenaillet) viennent reposer sur
le domaine brianonnais de la
marge europenne.
DES TMOINS
OCANIQUES
Certaines rgions des Alpes pr-
sentent des affleurements de
roches sdimentaires riches en
fossiles marins, tmoins de lan-
cien domaine ocanique :
lescalcairesrudistes(mollusques
bivalves) ouorbitolines (animaux
unicellulaires) des massifs du Ver-
corset delaChartreusetmoignent
de rcifs coralliens dans une mer
peuprofonde et chaude ;
les radiolarites du Chenaillet
sont formes par laccumulation
de squelettes en silice de radio-
laires, des animauxplanctoniques
unicellulaires ; elles tmoignent
dunocande grande profondeur.
ZOOM SUR...
LE DIAGRAMME
DE STABILIT
DES MINRAUX
DU MTAMORPHISME
Il sagit dun graphique qui per-
met de visualiser dans quelles
conditions de pression (donc de
profondeur) et de temprature les
minraux peuvent exister.
Les domaines de stabilit des
diffrents assemblages de mi-
nraux sont dtermins expri-
mentalement en laboratoire, en
les soumettant des pressions
et des tempratures croissantes
dans des presses appeles cel-
lules enclumes de diamant .
Ces outils permettent de couvrir
pratiquement tout le domaine de
pressions et de tempratures que
lon rencontre lintrieur de la
Terre.
Le diagramme peut tre dcoup
en facis mtamorphiques
(schistes verts, schistes bleus,
clogites, etc.) qui nindiquent
pas la nature de la roche, mais
les conditions de pression et de
temprature qui font apparatre
des associations minrales carac-
tristiques.
Ainsi, les minraux du mtamor-
phisme observs dans une roche
permettent de reconstituer les
conditions auxquelles cette roche
atsoumiseaucours delhistoire
de la chane de montagnes.
LE PRINCIPE
DES PRESSES
MINRAUX
Les cellules enclumes de dia-
mant permettent de soumettre
les minraux des pressions
allant jusqu 500 gigapascals
(GPa).
Une pression correspond une
force par unit de surface. Le prin-
cipe de cette technique repose
ainsi sur lapplicationdune force
trs importante sur la surface la
plus rduite possible. Le diamant
est utilis cette n depuis les
annes 1950 pour ses proprits
de rsistance exceptionnelles. En
effet, les diamants se forment
eux-mmes grande profondeur,
cest--dire des pressions trs
leves.
UN SUJET PAS PAS
Le domaine continental et sa dynamique Le domaine continental et sa dynamique
0,7mm
GT : grenat
Gl : glaucophane
J : jadite
GT
GI
J
J
100 200 300 400 500 600 700 800 Temprature(enC)
0,5
1
1,5
pyroxne
pagioclase
hornblende
pagaioclase
(glaucophane
grenat plagioclase)
plagioclase
grenat
grenat
jadite
quartz
glaucophane
glaucophane
glaucophane
jadite
quartz
grenat
jadite
quartz
plagioclase
plagioclase
actinote
chlorite
Remarque : les transformations minrales tant trs lentes,
des minraux forms dans un domaine de temprature et
pression donn peuvent tre encore prsents mme si la
roche nest plus dans ce domaine (minraux reliques).
Source : Centre Brianonnais de Gologie Alpine
Document 1 : Coupe gologique schmatique des
ophiolites duChenaillet
Cinf 1 et Cinf2 : couches dates du crtac infrieur.
Cinf 1 est une couche plus ancienne que Cinf2. Elle est
constituedpaissesmassesdecalcairesblancs Rudistes
et Orbitolinids (mollusques et foraminifres marins
fossiles).
Les basaltes suprieurs sont gs de 100millions dannes
Lesplusvieuxsdimentsocaniques(nongurssurlacoupe)
qui surmontent lesbasaltesont 65millionsdannes.Source:
Comprendreet enseigner laplanteterre. Caronet al. Ophrys
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
48
LES ARTICLES DU
49
LES ARTICLES DU
Pour les sismologues, ce
tremblement de terre nest
pas encore le big one
L
e sisme de Honshu
nest pas le big one
que les spcialistes
redoutaient dans la rgion
de Tokyo. Mais avec une ma-
gnitude de moment de 8,9,
selon le service gologique
amricain (USGS), cest lui
aussi un big one . Il sagit
de la secousse la plus puis-
sante quait connue le Japon
depuis que des rseaux de
sismomtres sont en place ,
indique Jrme Vergne, de
lInstitut de physique du
globe (IPG) de Strasbourg. Il
fait partie des dix plus gros
sismes enregistrs dans
le monde depuis un sicle,
indique le chercheur. Ce
sisme, qui nest pas une
surprise dans la rgion, a t
plus puissant que ce quoi
on sattendait , confirme son
collgue Pascal Bernard, de
lIPG de Paris.
Le tremblement de terre est
survenu prs de la cte est de
lle principale de larchipel,
130 km au large de la ville
de Sendai, par 25 km de pro-
fondeur. Il rsulte des mouve-
ments relatifs entre la plaque
nord-amricaine, qui englobe
une portion de lEurasie, et la
plaque pacique, celle-ci glis-
sant vers louest une vitesse
de 83 millimtres par an la
latitude du sisme. Le m-
canisme au foyer sisme, que
nous commenons analyser,
est dit en faille inverse : la
plaque Pacique plonge cet
endroit sous lEurasie , in-
dique Jrme Vergne.
Cette rupture a engendr
une srie de vagues parce
quelle est intervenue prs
de la surface et a entran un
mouvement de masse deau
important. L encore, des
analyses sont en cours pour
dterminer lampleur du d-
placement vertical, probable-
ment de quelques mtres, qui
pourra tre prcise laide
dimages radar du relief de lle
de Honshu.
Le sisme a t prcd par une
srie de gros tremblements de
terre : le 9 mars, une secousse
dune magnitude de 7,2 avait
eu lieu 40 km environ, sui-
vie de trois autres vnements
dune magnitude suprieure
6 le mme jour.
La zone de subduction de la
fosse du Japon a t le thtre
de neuf vnements dune
magnitude suprieure 7 de-
puis 1973, rappelle lUSGS. Le
plus gros tait un sisme de
magnitude 7,8 survenu en d-
cembre 1994 230 km au nord
du sisme de Honshu. Il avait
fait trois morts et 700 blesss.
Les sismologues vont se
concentrer sur lvaluation de
ltendue de la fracture. Il est
important de dterminer la
longueur de cette faille, car
cela nous donnera des indica-
tions sur lampleur possible
des rpliques , indique Pascal
Bernard, qui nexclut pas des
secousses dune magnitude de
7,5 8, accompagnes de
risques de tsunami impor-
tants. Le chercheur sinquite
aussi de limpact du sisme de
Honshu sur la zone tectonique
plus proche de Tokyo, o le
big one tait redout. La
menace est dautant plus
grande que la zone a t per-
turbe. Cest cela que nous al-
lons dsormais tudier.
Herv Morin
(12 mars 2011)
POURQUOI
CET ARTICLE ?
Lactualit rcente au Japon
met en exergue limportante
sismicit lie auxzones de sub-
duction. Dans ce cas, il sagit de
la plaque Pacique qui plonge
sous lEurasie avec une vitesse
de plus de 8 cm/an. Larticle
montreainsi lancessitdune
connaissance approfondie des
structures gologiques et de la
dynamique des plaques pour
la protection des populations
vivant dans les zones actives
du globe.
Des sismes provoqus
par le ballet des continents
Alaska, Kamtchatka, Chili, Asie du Sud-Est et Pacique sont situs sur des zones de
subduction lorigine des plus violentes secousses.
L
es sismes destructeurs
qui ont frapp Sumatra
en 2004 et 2005, et au-
jourdhui Java, rsultent des
mouvements lents et puissants
de la crote terrestre. Celle-ci
est forme dune douzaine de
plaques qui scartent les unes
des autres au fond des ocans,
sous leffet des courants de
convection du manteau ter-
restre, un peu comme ceux qui
animent leau qui bout dans
une casserole. Ce mcanisme
permet la crationdune crote
neuve qui repousse les plaques
existantes et fait plonger celles
qui sont ocaniques, donc plus
lourdes, sous dautres, plus l-
gres, engnral continentales.
Ce processus dit de subduc-
tion se produit au niveau
des grandes fosses ocaniques,
et cest lui qui est luvre
en Indonsie, o la plaque
indo-australienne ocanique
plonge sous la plaque eurasia-
tique continentale au rythme
de 5 6 cm par an. Cela peut
paratre faible, mais cest une
vitesse considrable eu gard
aux forces titanesques mises
en jeu dans ce mouvement de
plonge.
Outre la production de sismes
violents, la subduction donne
aussi lieu un volcanisme im-
portant. Car lorsquune plaque
plonge sous une autre, il se
produit, vers 100 km de pro-
fondeur, un changement dans
la structure des minraux qui
saccompagne de chaleur et fait
fondre partiellement la zone de
subduction. Ce matriel chaud
et moins dense a tendance
remonter. Et par unmcanisme
trs complexe, il peut donner
lieu des ruptions.
Bien dautres rgions du globe
sont confrontes ce phno-
mne de subduction lori-
gine dune trs forte activit
sismique. Celle-ci nest pas
systmatiquement meurtrire
car nombre de ces zones de
subduction sont dans des
rgions quasi dsertiques ou
peu peuples , rappelle Paul
Tapponnier, spcialiste de tec-
tonique lInstitut de physique
du globe de Paris.
Cest le cas de lAlaska, qui
connat rgulirement de trs
gros sismes, comme celui de
magnitude 9,2 qui la secou en
1964. Cest celui duKamtchatka,
qui en a enregistr un de ma-
gnitude 9 en 1952. Le Chili nest
pas labri en raison de la plon-
ge de la plaque Nazca sous
lAmrique du Sud. Situation
qui a donn lieu en 1960 au
plus gros sisme de lhistoire
rcente avec une magnitude re-
cord de 9,5. Et dautres de cette
importance sont attendus par
les sismologues, dans une zone,
esprent-ils, quasi dsertique
de la cte.
Les les Tonga-Kermadec et les
les Mariannes dans le Pacique
sont galement sous la menace
de telles secousses. Mais ces
rgions sont faiblement peu-
ples, au contraire du Japon,
des Philippines, de Tawan, de
lIndonsie, des Antilles ou de
la Mditerrane orientale. Sy
ajoutent, pour certains de ces
pays, des normes de construc-
tion parasismique peu contrai-
gnantes, voire inexistantes.
Les sismes de subduction les
plus importants (interplaques)
sont provoqus par le frotte-
ment entre deux plaques tec-
toniques. Mais cela ne suft
pas. Il faut aussi que la zone
de contact soit trs longue,
large et dune pente trs faible,
explique Louis Dorbath, sismo-
logue lInstitut de physique
du globe de Strasbourg. Cest
ce qui sest pass au Chili et
Sumatra, par exemple, mais pas
aux les Tonga et Mariannes,
o la pente est trs forte et
linterface, faible.
La plus grande partie de lner-
gie sismique du globe est dis-
sipe dans les zones de subduc-
tion. Mais tous les grands
sismes terrestres ne sont pas
dus ce mcanisme. Il existe
aussi des rgions de collision
continentale frontale qui don-
nent lieu des secousses tellu-
riques, comme celle qui a
produit lHimalaya lorsque
lancien ocan Thtys a t
entirement ferm par lavan-
ce de la plaque indienne
subductant sous la plaque
eurasiatique. La marge conti-
nentale de lInde est alors
arrive au contact de lAsie,
et la formidable collision
des continents a commenc.
Ctait il y a cinquante mil-
lions dannes.
Christiane Galus
(31 mai 2006)
POURQUOI
CET ARTICLE ?
Cet article rsume les origines
delactivitsismiqueet dumag-
matisme caractristiques des
zones de subduction. Attention
toutefois aux raccourcis voca-
teurs concernant la convection
dans le manteau terrestre, ils
sont manipuler avec prcau-
tion. De mme, les programmes
ofciels insistent davantage sur
laugmentation de la densit de
lalithosphreocaniquecomme
origine de la subduction.
Le domaine continental et sa dynamique Le domaine continental et sa dynamique
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
50
LESSENTIEL DU COURS
Le domaine continental et sa dynamique 51
LESSENTIEL DU COURS
Le domaine continental et sa dynamique
Des zones daccrtion
continentale
Les roches issues du magmatisme des zones de
subduction peuvent prsenter des compositions
chimiques proches, mais sont de deux types :
des andsites, roches associes au volcanisme
explosif, qui contiennent des phnocristaux, dont
de nombreux feldspaths plagioclases ;
des granodiorites, roches plutoniques formes
de quartz, feldspaths plagioclases, micas et amphi-
boles, faisant partie des granitodes.
Une grande partie des magmas cristallisent en
profondeur sous la forme de granitodes, que
lrosion nit par mettre jour. Ces roches forment
donc de la nouvelle crote continentale : on parle
daccrtion continentale.
Lorigine des magmas
Ltude de la compositionchimique des granitodes
montre quils sont issus du refroidissement dun
magma dorigine mantellique. Cela suggre une
fusionpartielle des pridotites laplomb des zones
dactivit volcanique, cest--dire dans le manteau
de la plaque chevauchante.
Or les conditions de pression et de temprature
qui rgnent au niveau des zones de subduction ne
permettent pas denvisager une fusionpartielle des
pridotites (le solidus ne recoupe pas le gotherme),
moins que celles-ci ne soient hydrates.
Lorigine de leau
Depuis sa formation au niveau de la dorsale, la
lithosphre ocanique qui entre ensubductionsest
hydrate : elle est riche en minraux hydroxyls
(OH
). Entranes en profondeur, les roches subis-
sent un mtamorphisme de haute pression mais
basse temprature, la subduction tant plus rapide
que le rchauffement de la lithosphre.
Les minraux caractristiques de la subduction
(glaucophane, jadite, grenat) sont moins riches en
eau : les ractions caractristiques du mtamor-
phisme de haute pression / basse temprature
entranent une libration deau. Elle percole dans
le manteau de la plaque chevauchante et abaisse la
temprature de fusion des pridotites. Entre 80 et
180 km de profondeur, le solidus humide croise
le gotherme de subduction : il y a fusion partielle
et production de magma, lorigine des andsites
et granitodes.
Le magmatisme en zone
de subduction : une
production de nouveaux
matriaux continentaux
L
es ocans peuvent tre bords par des marges actives, corres-
pondant des zones au niveau desquelles la lithosphre oca-
nique plonge dans lasthnosphre. Les zones de subduction
sont le sige dune importante activit magmatique qui aboutit
la production de crote continentale par des mcanismes lis aux
transformations minralogiques de la plaque plongeante.
Unelamemincedandsiteobserveaumicroscopeenlumire
polarise-analyse
1. feldspathplagioclase/2. amphibole/3. ptemicrolitique
0
0
25.10
3
50.10
3
75.10
3
80
160
240
Profondeur
(km)
Pression
(hPa)*
* hPa : hectopascals
SOLIDE
SOLIDE
+
LIQUIDE
LIQUIDE
1 000 2 000 T (C)
Diagramme pression/temprature, daprs Comprendre et
enseigner laplanteTerre, Caron-Ophrys
Trac des isothermes
en zone de subduction
Le mtamorphisme caractristique des zones de
subduction est dit de haute pression et basse
temprature (HP-BT). Il ne sagit pourtant pas de
basses tempratures proprement dites, mais de
tempratures plus basses queneleprvoit legradient
gothermique habituel, cest--dire lvolution de
la temprature en fonction de la profondeur dans
le globe. En effet, la lithosphre ocanique froide
senfonce dans lasthnosphre trop rapidement
pour que lquilibre thermique se ralise. Le trac
des isothermes permet ainsi de visualiser la plaque
plongeante.
Arc magmatique Fosse
Crote
200c
400c
600c
800c
1000c
1200c
200c
400c
600c
800c
1000c
1200c
Crote
Profondeur
L
ith
o
sp
h
re
Modle en coupe de trac des isothermes (lignes dgale
temprature) dans une zone de subduction
MOTS CLS
ACCRTION
CONTINENTALE
Formation de nouvelle crote
continentale au niveau des zones
de subduction. Les roches se for-
ment par cristallisationdumagma,
issu de la fusion partielle des pri-
dotiteshydratesdumanteausitu
au-dessus delaplaqueplongeante.
SUBDUCTION
Plongement dune lithosphre
ocanique dans lasthnosphre
situe sous une autre plaque, elle-
mme constitue de lithosphre
ocanique ou continentale. Une
zone de subduction constitue une
limitedeplaquecaractrisepar la
prsence dune fosse ocanique et
uneactivitsismiqueetvolcanique.
ZOOM SUR
Lactivitsismiquedeszonesde
subduction
Au niveau des zones de subduc-
tion, des sismes foyers profonds
sont enregistrs (jusqu plusieurs
centaines de kilomtres de pro-
fondeur). Les foyers des sismes
sont aligns le long du plan de
Benioff, qui matrialise la plaque
plongeante.
NOTIONS CLS
ROCHES MAGMATIQUES
Les roches plutoniques comme la
granodiorite et le granite provien-
nent du refroidissement lent du
magma en profondeur (pendant
plusieurs dizaines de milliers dan-
nes). Les minraux ont le temps
de cristalliser : la structure est dite
grenue(grainsvisibleslil nu).
Les roches volcaniques, comme
landsite, proviennent du re-
froidissement rapide du magma
la surface. La structure est dite
microlitique (ou vitreux) : des
cristaux sont noys dans un verre,
qui correspondaumagmadont les
atomes nont pas eu le temps de
sordonner encristaux.
PHNOCRISTAUX
Dsignent dans une roche volca-
nique les cristaux de grande taille,
visibles lil nu.
ZOOM SUR...
LVOLUTION
DE LA LITHOSPHRE
OCANIQUE
Au cours de lexpansion oca-
nique, la dformation est
cassante, la crote ocanique
se fracture. Les fracturations
permettent leau de mer trs
froide de pntrer.
Elle ressort haute temprature
(350 C) au niveau de chemines
hydrothermales (fumeurs noirs).
On parle de circulation hydro-
thermale .
Le flux thermique de la lithos-
phre diminue avec lge : en
sloignant de laxe de la dor-
sale, la lithosphre ocanique
refroidit.
La diminution de la temprature
et la circulation deau sont
lorigine de changements mi-
nralogiques au sein des roches
de la lithosphre ocanique. Les
nouveaux minraux sont hy-
drats : ils possdent dans leur
formule chimique des radicaux
hydroxyles OH
. Ils sont stables
basse temprature.
la place des feldspaths plagio-
clases et pyroxnes du basalte
et du gabbro, on trouve des
amphiboles, de la chlorite et de
lactinote (facis schistes verts).
Les pridotites sont serpentini-
ses : on constate lapparition de
la serpentine, au dtriment de
lolivine et des pyroxnes.
Il sagit dun mtamorphisme
basse pression / basse tempra-
ture (BP-BT).
Cest donc une lithosphre oca-
nique froide, dense et hydrate
qui entre en subduction.
LES RACTIONS DU
MTAMORPHISME HP-BT
AU COURS DE LA
SUBDUCTION
(1) plagioclase + chlorite* + ac-
tinote* glaucophane + eau
(facis schiste bleu)
(2) plagioclase + glaucophane
grenat + jadite + eau (facis
clogite)
* Chlorite et actinote sont des
minraux hydrats.
UN ARTICLE DU MONDE CONSULTER
Un strato-volcan gris situ aux franges de la plaque carabe p. 54
(Herv Morin, 20aot 1997)
Fosseocanique
Ocan
Croteocanique
Crotecontinentale
Andsite, rhyolite
L
ith
o
s
p
h
r
e
c
o
n
tin
e
n
ta
le
L
ith
o
s
p
h
r
e
o
c
a
n
iq
u
e
M
a
n
te
a
u
a
s
th
n
o
s
p
h
r
iq
u
e
Plutonde
granodiorite
Manteaulithosphrique
Subduction
S
ch
istes b
leu
s
c
lo
g
ite
s
Dshydratation
Sismes
Chambremagmatique
formationdumagma
andsitiquepar fusion
partielle
Schma bilan de l'origine du magmatisme en zone de subduction
Lorigine du magmatisme
en zone de subduction
La lithosphre ocanique qui entre en subduction
est ge, donc froide et paisse et, par consquent,
dense. Les roches contiennent des minraux hy-
drats. Au cours de lenfoncement de la lithosphre
ocaniquedans lasthnosphremoins dense, les mi-
nraux sont soumis une pression croissante et se
transforment en des minraux caractristiques des
conditions de haute pression/ basse temprature, en
particulier le grenat (facis des clogites). Au cours
de ces ractions mtamorphiques, les minraux
deviennent anhydres et leaulibre percole dans le
manteau de la plaque chevauchante. Lhydratation
des pridotites permet leur fusion partielle et le
magma ainsi produit alimente les volcans des zones
de subduction et forme des plutons de granitodes
en profondeur.
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
52 Le domaine continental et sa dynamique 53 Le domaine continental et sa dynamique
Les documents
Document 1 : Compositionminralogique dungab-
bro en fonction des conditions de pression et de
temprature
Enlaboratoire, onsoumet deschantillonsdegabbros
des conditions de pression et de temprature va-
riables. Les rsultats sont prsents sur le graphique.
Onrappellequelegabbro, lorsquil seforme, contient
presque exclusivement des plagioclases et du py-
roxne.
Deuxractions dumtamorphisme engendres par
une augmentation de pression :
Plagioclase+chlorite*+actinote*->glaucophane+eau
Plagioclase +glaucophane ->grenat +jadite +eau
* Chlorite et actinote sont des minraux hydrats.
Document 2 : Densits de quelques roches rencon-
tres dans une zone de subduction
Partie 2.1 : Mcanismes en jeu
dans les zones de subduction
Plaquesus-jacente
Magma Volcanisme
500C
0
15
30
60
Profondeur
enkm
600C
800C
1000C
1200C
1200C
Plaqueplongeante
Manteausuprieur
L
ith
o
s
p
h
re
o
c
a
n
iq
u
e
SV
SB
E
Crote ocanique
H
2 0
H
2 0
PointA
Manteau
Onprcisequelemanteau
est constitudepridotites
et lacroteocanique,
debasalteset degabbros.
Lgende:
Zonedefusionpartielle
delapridotitehydrate
SV Facisschistesverts
SB Facisschistesbleus
E Facisclogites
AUTRES SUJETS POSSIBLES AU BAC SUR CE THME
Partie 1, synthse sans documents :
Montrer comment latransformationdesroches, qui survient lorsdelaplongedelalithosphreocanique, est
lorigine dumagmatisme caractristique des zones de subduction. Schmas attendus.
Partie 2.1 :
Exploiter des documents pour orienter une subduction.
Partie 2.2 :
Exploiter des documents pour montrer quune rgioncorrespondbien une zone de subduction.
ZOOM SUR
Ladcouverte des zones
de subduction
Plusieurs annes avant llabora-
tion du modle de la tectonique
des plaques par le Franais Xavier
Le Pichon et ses collaborateurs
(1968), le sismologue amricain
Hugo Benioff a redcouvert et
gnralis les travauxduJaponais
Wadati. Cedernier avait lisismes
et volcans japonais ladrivedes
continents ds 1935, alors que les
ides de Wegener taient encore
refuses. Benioff a constat que
dans ces rgions lasismicitet au
volcanisme parfois spectaculaires
les tremblements de terre taient
localiss sur un plan plongeant,
qui sera alors baptis plan de
Wadati-Benioff . Certains sismes
se localisent des profondeurs si
importantes quil ne devrait pas y
avoir de dformation cassante en
raison des tempratures leves
qui rendent les matriauxductiles.
Les tudes menes dans les annes
1950 ont montr que ces rgions
concidaient aussi avec des fosses
sous-marines trs profondes. Dans
les annes 1960, des chercheurs
amricains ont montrqueleplan
de Wadati-Benioff correspondait
une plaque plongeante de lithos-
phre ocanique froide et suf-
samment rigide pour se casser,
provoquant les sismes.
PERSONNAGES
CLS
Les diables des volcans
Maurice et Katia Krafft sont des
volcanologues franais, ns en
1946 et 1942, ayant uvr dans le
monde entier pour la prvention
des risques volcaniques auprs
des populations. Ils ont sans doute
contribu sauver des centaines
de milliers de vies humaines,
comme celles des 300 000 per-
sonnes ayant accept dvacuer la
zone duPinatubo auxPhilippines
aprs avoir vu un de leurs lms.
Ironie du sort, le volcan entra en
ruption quelques jours aprs la
mort du couple, emport par une
nue ardente sur le mont Unzen
au Japon le 3 juin 1991.
ZOOM SUR
LES RUPTIONS
EXPLOSIVES
Laccumulation dune forte pres-
siondans lachambremagmatique
dunvolcandont les magmas sont
visqueux peut dclencher une
ruption explosive. Les nues ar-
dentesqui peuvent enrsulter sont
trs destructrices, et le panache de
cendres peut atteindre plusieurs
dizaines de kilomtres daltitude.
QUELQUES RUPTIONS
MARQUANTES :
le Vsuve en lan 79 ;
leKrakatoaenIndonsieen1883;
la montagne Pele la Marti-
nique en 1902 ;
le mont Saint Helens aux tats-
Unis en 1980;
le Nevado del Ruiz en Colombie
en 1985 ;
le Pinatubo en 1991 aux Philip-
pines ;
le Merapi en 2010en Indonsie.
UNE VILLE RAYE
DE LA CARTE
Le 8 mai 1902 la Martinique,
la ville de Saint-Pierre disparat
en quelques minutes au cours de
lruption explosive de la mon-
tagne Pele. 28 000 victimes et
seulement 2 survivants : un lourd
bilan li la minimisation du
danger par les autorits, malgr
les signaux dalerte. Fumerolles,
explosions, tremblements deterre,
panaches devapeur et decendres
sont autant de manifestations t-
moignant de limminence dune
ruption. Le 7 mai, le volcan est
calme et une commission scien-
tique a dclar quil ny avait pas
de risque.
Le terme nue ardente a t
introduit suite lruption de la
montagne Pele.
Depuis, lasurveillancedes volcans,
en particulier ceux des Antilles
franaises, sest dveloppeautour
dobservatoires volcanologiques.
Mais dans diffrentes rgions du
globe, les recommandations de
mesures prventives des scienti-
ques se sont souvent heurtes
aux dcisions politiques et aux
intrts conomiques, avec des
consquencesparfoisdramatiques.
Document de rfrence : Coupe schmatique dans une zone de subduction
1. Unmtagabbrode lacrote ocanique prlev par
forage au point A (cf. document de rfrence), trs
loinde laxe de la dorsale, contient les assortiments
de minrauxsuivants :
a) plagioclases + chlorites + actinotes.
b) plagioclases + glaucophanes.
c) glaucophanes + jadites.
d) grenats + jadites + glaucophanes.
2. Unmtagabbro plagioclase et glaucophane sou-
mis une augmentationde pression, temprature
constante, acquiert de la:
a) jadite et senrichit en eau.
b) jadite et libre de leau.
c) chlorite et libre de leau.
d) chlorite et senrichit en eau.
3. On considre les roches suivantes : (A) mtagab-
bro chlorite et actinote ; (B) gabbro plagioclases
et pyroxnes ; (C) mtagabbro glaucophane et
plagioclases ; (D) mtagabbro grenats et jadite ;
(E) pridotite.
Le classement de ces roches par ordre de densits
croissantes est :
a) E < A< B < C <D. b) B < A< C < D<E.
c) E < B < A< C < D. d) B < E < A< C < D.
4. Le point partir duquel la lithosphre ocanique
peut commencer senfoncer dans lasthnosphre
peut tre tabli encomparant :
a) lpaisseur de la crote et celle du manteau.
b) lge de la crote et celui du manteau.
c) la masse dune colonne de lithosphre et celle
dunecolonnedasthnosphre(demmesurface).
d) lpaisseur de la lithosphre et celle de lasth-
nosphre.
5. Lge, exprimenmillionsdannes, partirduquel
la lithosphre ocanique disparat en senfonant
dans lasthnosphre est :
a) 10. b) 25. c) 30. d) 80.
Analyse du sujet
Il sagit dexploiter des documents et de les mettre
en relation an de choisir une proposition exacte
parmi plusieurs. Le format QCMpeut paratre plus
facile, mais il a le dsavantage de ne pas permettre
de nuancer la rponse.
Le corrig
1. a), 2. b), 3. d), 4. c), 5. c.
Roches Densit
2,85
3,3
3,4
3,5
3,25
Lithosphre
Asthnosphre
Basaltes, gabbros
Mtagabbro en facis schistes verts
Mtagabbro en facis schistes bleus
clogites
Pridotites
Lintitul complet du sujet
On sintresse aux compositions minralogiques des roches prsentes dans une zone de subduction et au
moteur de la subduction (document de rfrence). On cherche comprendre certains mcanismes en jeu
dans les zones de subduction en exploitant les donnes prsentes dans les documents suivants. Cochez la
proposition exacte pour chaque question de 1 6.
25
Profondeur
(km)
75
50
Pression
en GPa
1
2
Temprature
en C
200 400 600 800
chlorite + actinote
+ plagioclase
glaucophane
+ plagioclase
glaucophane
+ jadite
grenat de haut
pression + jadeite
+/ glaucophane
ge de la lithosphre (en 10
6
ans)
Distance laxe de la dorsale
( )
(en km)
paisseur de la
lithosphre
ocanique (en km)
Crote
Manteau
Masse dune colonne de
lithosphre ocanique de surface
gale 1 m
2
(en 10
3
tonnes)
Masse dune colonne
dasthnosphre de mme
surface et de mme paisseur
(en 10
3
tonnes)
2 10 15 25 30 40 60 80 100
8 000 6 400 4 800 3 200 2 400 2 000 1 200 800 160
5 5 5 5 5 5 5 5 5
8 24 31 41 45 53 66 77 87
40,7 93,5 116,6 149,5 162,8 189,2 232,1 268,4 301,4
42,3 94,3 117,0 149,5 162,5 188,5 230,7 266,5 299,0
Les masses de la lithosphre ocanique et de lasthnosphre sont tablies pour une colonne de surface gale 1 m
2
.
Ce quil ne faut pas faire
Penser quon est incapable de rpondre car on
ne connat pas par cur tous les minraux du
mtamorphisme (il faut utiliser les documents et la
logique pour procder par limination).
Utiliser les documents sans les mettre en relation.
Se dispenser de brouillon pour rpondre aux
questions dont la rponse nest pas immdiate.
Document 3 : Quelques caractristiques de lalithosphre et de lasthnosphre
UN SUJET PAS PAS UN SUJET PAS PAS
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
54
LES ARTICLES DU
ENJEUX PLANTAIRES
CONTEMPORAINS
130
120
110
100
90
80
70
Flux de chaleur
60
510' 0' ' N
80' 0' ' E 40' 0' ' W 20' 0' ' W 00' 0' ' E 20' 0' ' E 40' 0' ' E 60' 0' ' E
80' 0' ' E 40' 0' ' W 20' 0' ' W 00' 0' ' E 20' 0' ' E 40' 0' ' E 60' 0' ' E
490' 0' ' N
470' 0' ' N
450' 0' ' N
430' 0' ' N
510' 0' ' N
490' 0' ' N
470' 0' ' N
450' 0' ' N
430' 0' ' N
0 200KM
Flux de chaleur (cor r ig)
1. manteau infrieur
2. noyau
3. asthnosphre
4. lithosphre
point chaud
point
chaud
subduction et cordillre associe
dorsale
ocanique
1
2
3
4
POURQUOI
CET ARTICLE ?
Cet article sur le volcan Sou-
frire Hills de Montserrat
dcrit les caractristiques des
volcans des zones de subduc-
tion et de leurs ruptions. Il
permet de raliser que ldice
volcanique sest constitu au
cours du temps par lempile-
ment successif de coules de
lave et de projections de blocs
et de cendres, lies lactivit
magmatique. Larticle prcise
les risques associs ces vol-
cans gris , risques dautant plus
importants que leurs pentes
fertiles attirent des populations
humaines nombreuses.
Le domaine continental et sa dynamique
Le vulcanologue Maurice Krafft,
qui avec sa femme Katia a pri
dans l'explosiondumont Unzen
auJaponle 3 juin1991, les appe-
lait volcan gris . Parce qu'ils
mettent roches et nus, mais
peu de lave liquide. Le volcan
de Soufrire Hills de Montserrat
fait partie de ces volcans explo-
sifs, plus dangereux que leurs
cousins effusifs rouges qui,
Hawa par exemple, s'pan-
chent paisiblement en gerbes
colores.
Une autre typologie rattache
Soufrire Hills aux strato-vol-
cans stromboliens, famille qui
compte les redoutables monts
Saint Helens, Pinatubo ou en-
core mont Unzen. Le volcan de
Monserrat est cousindenombre
de soufrires (Guadeloupe,
Sainte-Lucie, Saint-Vincent),
sans que cette homonymie si-
gnie pour autant qu'ils sont
relis souterrainement.
Ces strato-volcans sont consti-
tus de couches successives de
coules de lave et de couches
pyroclastiques formes de pro-
jections de blocs, de lapilli et de
cendres. Ce type de volcans, aus-
si dit composites , se trouve
gnralement le longdes arcs de
subduction : Soufrire Hills est
issu, tout comme le chapelet des
huit autres volcans des Antilles,
de l'affrontement des plaques
Pacique et Atlantique, qui se
rapprochent de quelques centi-
mtres par an et plongent sous
la petite plaque Carabe, prise
entenaille. Auxfranges de celle-
ci, des poches magmatiques se
constituent en profondeur. Ces
roches visqueuses cherchent
s'vacuer en surface, contri-
buant au l des ruptions qui
se sont succd ces derniers
millions d'annes laformation
des les.
Les strato-volcans sont parti-
culirement redouts. D'abord,
parce qu'ils sont les plus nom-
breux (60 % du total) et que
la terre fertile attire gnra-
lement leur pied une vaste
population. Ensuite, parce que
l'empilement qui les caract-
rise est souvent fragilis par
les inltrations d'eau, les trem-
blements de terre incessants
et la pousse magmatique, qui
favorisent les glissements de
terrain, les coules de boue et
les avalanches. Ainsi, le anc
norddumont Saint Helens s'est-
il effondr durant les premires
phases de l'ruption de 1980.
En 1792, le mme phnomne
avait entran la formationd'un
raz-de-mare meurtrier (15 000
victimes) au pied du mont Un-
zen. Des coules de boue ont
fait 23 000 morts au pied du
Nevado del Ruiz, en Colombie,
le 13 novembre 1985. En 1991,
ces lahar qui ont dval les
pentes du Pinatubo aux Philip-
pines ont fait 200 victimes.
Montserrat risque-t-elle de
tels cataclysmes ? La probabi-
lit d'une destruction totale
de l'le, similaire celle cause
en Indonsie par l'explosion
du Krakatoa en 1883 (36 000
morts), parat inme. Si un tel
phnomne devait se produire,
il serait, selon les spcialistes,
annonc par des signes avant-
coureurs trs clairs. Par ailleurs,
le relief de l'le met sa partie
nord l'abri des coules pyro-
clastiques, qui seront guides
dans des valles dont la popula-
tion a t vacue. En revanche,
les cendres et les bombes
issues des explosions peuvent
retomber sur l'ensemble de
Monserrat.
Pour l'heure, le dernier com-
muniqu de l'Observatoire du
volcan de Montserrat (MVO)
indiquait, lundi 18 aot, que
l'activit de Soufrire Hills
reste d'un niveaulev et est
caractrise par des nues, des
secousses telluriques et des
chutes de pierres continues. De
fortes missions de vapeur ont
t observes sur le anc nord
dudme, et de nouvelles explo-
sions pourraient survenir
avec ou sans signes avant-
coureurs , se traduisant par la
chute de pierres et de cendres
sur toute l'le, tandis que des
coules pyroclastiques sont
attendues sur toutes les faces
du volcan. Aussi l'Observatoire
recommande-t-il la popula-
tion de se tenir prte s'abriter
tout moment sous des abris
ou des toitures solides, de dis-
poser d'un casque et de porter
des masques pour se protger
de la poussire.
Herv Morin
(20aot 1997)
Un strato-volcan gris
situ aux franges
de la plaque carabe
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
56
LESSENTIEL DU COURS
Enjeux plantaires contemporains 57
LESSENTIEL DU COURS
Enjeux plantaires contemporains
Gothermie et proprits
thermiques de la Terre
Flux et gradient gothermiques
la surface de la Terre, de nombreuses manifes-
tations tmoignent de lexistence de matriaux
chauds en profondeur : ruptions volcaniques,
sources chaudes hydrothermales, augmentation
de la temprature dans les mines, etc. Des sources
deau chaude, comme Chaudes-Aigues dans le
Massif central, sont utilises depuis lAntiquit, alors
que les premires mesures de ux gothermique
ont t ralises, en France, dans les annes 1960.
Le ux gothermique correspond au produit du
gradient gothermique vertical par la conductivit
thermique des roches. Le ux gothermique et le
gradient gothermique sont mesurs sur chaque
site dtude, tandis que la conductivit thermique
des roches est dtermine en laboratoire.
Les valeurs des gradients et des ux gothermiques
varient selon le contexte godynamique. En do-
maine ocanique, les uxgothermiques sont levs
au niveau des dorsales ocaniques, des points
chauds (le de La Runion, par
exemple) et des arcs volcaniques
ou cordillres lis la subduc-
tion (Japon, Andes, Antilles, etc.).
En domaine continental, les ux
gothermiques sont forts au ni-
veau des zones avec magmatisme
(valle du rift africain). De plus,
des bassins sdimentaires ayant
une crote amincie prsentent un
ux gothermique lev, tels les
bassins deffondrement de lAlsace
oude la Limagne enFrance. Quelle
est lorigine de cette nergie ther-
mique observable en surface ?
Lorigine de lnergie
gothermique et les
transferts thermiques
dans la Terre
Lnergie gothermique interne
provient essentiellement de la
dsintgration des lments ra-
dioactifs (uranium :
238
U et
235
U ;
L
a grande majorit de lnergie actuellement utilise (agricul-
ture, industrie, transport, etc.) provient soit de la combus-
tion dnergies fossiles, comme le ptrole ou le charbon, soit
du nuclaire. Or, les nergies fossiles ne sont pas renouvelables
et pourraient manquer dans lavenir. De plus, leur combustion, en
faisant augmenter le niveau de CO2 dans latmosphre, est lori-
gine dun effet de serre entranant une augmentation de la tem-
prature terrestre. Avec lnergie nuclaire se posent le problme
de sa sret et celui de la gestion des dchets long terme. Le
dveloppement durable sintresse dautres lires nerg-
tiques, plus respectueuses de lenvironnement et quasiment in-
puisables pour lhomme, comme la gothermie. O et comment
est exploite lnergie gothermique et quelle est son origine ?
130
120
110
100
90
80
70
Flux de chaleur
60
510' 0' ' N
80' 0' ' E 40' 0' ' W 20' 0' ' W 00' 0' ' E 20' 0' ' E 40' 0' ' E 60' 0' ' E
80' 0' ' E 40' 0' ' W 20' 0' ' W 00' 0' ' E 20' 0' ' E 40' 0' ' E 60' 0' ' E
490' 0' ' N
470' 0' ' N
450' 0' ' N
430' 0' ' N
510' 0' ' N
490' 0' ' N
470' 0' ' N
450' 0' ' N
430' 0' ' N
0 200KM
Flux de chaleur (cor r ig)
Le ux gothermique en France (obtenu partir de 479 mesures et complt par
desmesuresenmer et danslespayslimitrophes, daprsLucazeauetVasseur, 1989)
thorium :
232
Th ; potassium :
40
K) prsents dans les
roches des diffrentes enveloppes du globe.
Lnergie thermique produite dans la Terre est
transfre dans le globe selon deux modalits : la
conductionet la convection. Lefcacit dutransfert
thermique de la conduction dpend de la conducti-
vit thermique des matriaux.
Ainsi auseinde la lithosphre et linterface noyau/
manteau, lnergie thermique est transfre par
conduction. Par exemple, dans la crote terrestre,
le gradient gothermique moyen est de 30 C.km
1
.
La convection se met en place lorsquun matriau
chaud et peu dense est situ sous un matriau
plus froid et plus dense. Le matriau chaud et
peu dense slve, tandis que le matriau froid et
plus dense descend et se rchauffe son tour. Ces
mouvements de matire constituent des cellules
de convection qui existent dans le manteau et le
noyau. Le manteauest ainsi anim de mouvements
lents de convection, qui entranent le dplacement
des plaques lithosphriques situes au-dessus.
Le fort ux gothermique dans les dorsales sex-
plique par la remonte de matriel chaud
et la productionde lithosphre ocanique
nouvelle. Dans les zones de subduction,
la faible valeur du flux gothermique
moyen sexplique par le plongement de
la lithosphre ocanique ge, froide et
dense. Localement, dans les zones de sub-
duction, le magmatisme li la remonte
de matriel chaud est lorigine dun
ux thermique lev au niveau des arcs
volcaniques ou des cordillres. Ainsi, la
Terre est une machine thermique, dont la
dissipation de lnergie interne constitue
le moteur de la dynamique des plaques
lithosphriques.
La gothermie : une source dnergie
au service du dveloppement durable
Lnergie gothermique utilise actuellement par
lhomme couvre environ 1 % des besoins nerg-
tiques mondiaux pour la production dlectricit
et la production de chaleur. Le principe de la
production de chaleur par gothermie consiste
en la rcupration de lnergie thermique deaux
profondes, naturellement prsentes ou apportes
par des conduites en profondeur. travers le
monde, environ 350 centrales gothermiques
produisent de llectricit. Ces centrales gother-
miques sont en majorit implantes dans des
zones de subduction (Japon, Nouvelle-Zlande,
Antilles, etc.). Dautres centrales sont situes au
niveau de dorsales ocaniques (Islande, Aores), de
fosss deffondrement (Alsace, Kenya) ou de points
chauds (Islande, Hawa), mais galement dans
des rgions stables du globe, comme les bassins
sdimentaires (tats-Unis, Chine, etc.).
Le prlvement de lnergie gothermique par
lhomme ne reprsente quune infime partie de
lnergie thermique dissipe la surface de la
Terre. La gothermie constitue une
source dnergie inpuisable
lchelle humaine, propre, colo-
gique et conomique. La gothermie
est une source dnergie au service
du dveloppement durable. La di-
versit des implantations gother-
miques actuelles dans le monde
montre limportance de cette source
dnergie, que lhomme pourrait
utiliser davantage quil ne le fait
actuellement.
1. manteau infrieur
2. noyau
3. asthnosphre
4. lithosphre
point chaud
point
chaud
subduction et cordillre associe
dorsale
ocanique
1
2 3
4
Laconvectiondanslemanteau(modledeuxtagesdecellulesdeconvection)
UN ARTICLE DU MONDE
CONSULTER
Lthiopie se rve en futur
gant de la gothermie p. 59
(Pierre Lepidi, 21 aot 2011)
CHIFFRES CLS
En 2010, la France a consomm
266 Mtep (1 Mtep = 1 million de
tonnes d'quivalent ptrole, soit
lnergie dgage par un million
de tonnes de ptrole). En qua-
rante ans, la consommationner-
gtique de la France a presque
doubl mais se stabilise depuis
quelques annes. Ses besoins
nergtiques sont couverts par
le ptrole (30,9 %), le charbon
(4,1 %), le gaz naturel (15,0 %),
llectricit dorigine nuclaire
(41,8 %) et lensemble des ner-
gies renouvelables (8,2 %, nergies
hydraulique, olienne, photovol-
taque, gothermique, issue de la
valorisation des dchets, etc).
MOTS CLS
CONDUCTION
Transfert dnergie thermique de
proche en proche sans dplace-
ment de matire.
CONVECTION
Transfert dnergie thermique
avec dplacement de matire et
qui constitue un mcanisme de
transfert dnergie efcace.
FLUX GOTHERMIQUE
Quantit dnergie thermique
dissipe par unit de surface
terrestre et par unit de temps
(unit : W.m
2
). Le ux gother-
mique moyen est de lordre de
87 mW.m
2
. En gnral, il est
plus faible sur les continents
(~ 65 mW.m
2
) que sur les ocans
(~ 100 mW.m
2
).
GRADIENT THERMIQUE
Variation de temprature entre
deux points de lenveloppe ter-
restre rapporte la distance les
sparant (C.m
-1
).
MACHINE THERMIQUE
La Terre est une machine ther-
mique car dans le globe terrestre
ont lieu des transferts dnergie
thermique entre deux sources
d'nergie thermique dont les
tempratures sont diffrentes :
lintrieur de la Terre (source
chaude) et lespace qui lentoure
(source froide).
ZOOM SUR...
LE GRADIENT GO-
THERMIQUE EN FRANCE
Le gradient gothermique peut
tre compar endiffrents points
du territoire franais : 1,5 km de
profondeur, la temprature est de
70Cdans le Bassinparisien, alors
quelle est de 100 C en Alsace.
Dans les Antilles, au niveau de
lle de la Guadeloupe, la tem-
prature est de 250 C 1 km de
profondeur. Cette valeur leve
sexplique par la subduction af-
fectant cette rgion.
LES EXPLOITATIONS GO-
THERMIQUES EN FRANCE
La seule centrale gothermique
de France mtropolitaine, mise en
service depuis 2008, est installe
Soultz-sous-Forts, enAlsace, au
niveau du foss rhnan. En effet,
le foss rhnan prsente le plus
fort gradient gothermique de la
mtropole et est donc propice
linstallation dune centrale go-
thermique. Cettecentralegother-
mique utilise la technique de la
gothermie des roches fractures :
de leau froide est injecte plus
de 5 000 m de profondeur dans
des roches fractures. Cette eause
rchauffe au contact des roches.
Leau chaude est alors extraite
une temprature de 200 C et est
utilise pour alimenter des tur-
bines produisant de llectricit.
La Guadeloupe, situe dans une
zone de subduction, prsente un
fort ux gothermique. la cen-
trale gothermique de Bouillante,
de la vapeur deau 250 C jaillit
directement du forage et est utili-
se pour alimenter des turbines et
produire de llectricit.
Lexploitationgothermiquebasse
energie, servant notamment
chauffer des btiments, est locali-
se dans les bassins sdimentaires
(le-de-France, Aquitaine).
Actuellement lagothermierepr-
sente environ1 %de la couverture
nergtiquefranaise, mais sapart
dans lapprovisionnement nerg-
tique franais devrait augmenter
dans les annes venir, commeles
autres nergies renouvelables que
sont lolienterrestre et enmer, et
le photovoltaque.
Le ux gothermique mondial
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
Enjeux plantaires contemporains 58
LESSENTIEL DU COURS UN SUJET PAS PAS
Enjeux plantaires contemporains 59
Lintitul complet du sujet
Cochez la proposition exacte pour chaque question
de 1 6.
1. Les zones noires et engris visibles sur lacarte
reprsentent :
a) un ux thermique lev.
b) un ux thermique faible.
c) des diffrences de temprature ocanique.
d) des courants ocaniques chauds.
2. Les zones de uxfort visibles sur lacarte :
a) sont lies une plus forte nergie solaire arrivant
la surface de la Terre.
b) sont lies un volcanisme de point chaud.
c) sont en relation avec des dorsales.
d) sont lies la prsence des ocans la surface
du globe.
3. Les zones de uxfort sont associes :
a) aux zones de subduction.
b) la cration dasthnosphre.
c) la cration de lithosphre ocanique.
d) la cration de lithosphre continentale.
4. Lnergie gothermique exploitable par lhomme :
a) est constante dune rgion lautre.
b) est maximale au niveau de la lithosphre
continentale.
c) est optimale en Islande au niveau dun rift.
d) contribue fortement la couverture nergtique
de lhumanit.
Le document ci-dessous reprsente le ux thermique
au niveau du plancher ocanique en fonction de son
ge en millions dannes. Chaque bote reprsente la
variabilitdesdonnesdanslatranchedgeconsidre.
5. Le graphique montre que :
a) le ux thermique du plancher ocanique
augmente lors de son vieillissement.
b) le plancher ocanique senfonce au fur et
mesure de son vieillissement.
c) le plancher ocanique se refroidit en vieillissant.
d) le ux thermique est constant au niveau du
plancher ocanique.
6. Onpeut associer les zones duplancher ocanique
uxthermique faible avec :
a) une lithosphre sous-jacente plus ne et plus
dense.
b) une lithosphre sous-jacente plus paisse et plus
lgre.
c) une lithosphre sous-jacente plus ne et plus
lgre.
d) une lithosphre sous-jacente plus paisse et plus
dense
Le corrig
1. a), 2. c), 3. c), 4. c), 5. c), 6. d)
Partie 1 : Lnergie gothermique
mWm-2
Les lignes noires reprsentent les contours
continentaux et les limites de plaque
0 85 120 180 350
Carte mondiale des ux thermiques exprims en
milliwattspar mtrecarr(mW.m
2
)
400
300
200
100
0
0 40 80 120 160 200
ge (Ma)
.
Ce quil ne faut pas faire
Cocher deux rponses ou ne cocher
aucune rponse.
Ngliger les documents quand leur utilisation est
ncessaire pour rpondre la question.
AUTRES SUJETS POSSIBLES AU BAC SUR CE THME
Matrise des connaissances avec questionde synthse :
Envousappuyant sur quelquesexemples, montrezqueleuxet legradient gothermiquesvarient enfonction
ducontexte godynamique.
tude de documents :
tude dune rgion : relations entre ux et gradient gothermiques, donnes de la tomographie sismique,
contexte godynamique et possibilit dexploitationgothermique.
ZOOM SUR...
LA TOMOGRAPHIE
SISMIQUE
Il sagit dune technique per-
mettant de cartographier lint-
rieur de la Terre en tudiant la
vitesse des ondes sismiques. La
tomographie vise corrler les
anomalies de vitesse des ondes
sismiques avec des variations
de composition chimique, dtat
physique ou de temprature
des diffrents milieux compo-
sant les enveloppes de la Terre.
Une anomalie dite ngative
correspond, dans une rgion
donne, une diminution de la
vitesse des ondes sismiques.
linverse, une anomalie posi-
tive correspond une augmen-
tation de la vitesse de ces ondes.
Dans une zone du globe terrestre
o la minralogie des roches est
identique, une anomalie nga-
tive indique une zone moins
dense, donc plus chaude. Une
anomalie positive est interpr-
te comme lexistence dune
zone plus dense, donc plus
froide. Ainsi la tomographie
sismique permet daccder
la connaissance des caractris-
tiques des enveloppes internes
de la Terre.
LA GOTHERMIE TRS
BASSE NERGIE
La gothermie domestique
dite trs basse nergie
utilise des sources dnergie
thermique dont la temp-
rature est de moins de 30 C.
Des pompes chaleur per-
mettent de puiser lnergie
thermique contenue dans les
couches superficielles du sous-
sol. Il est ainsi possible de
chauffer et de produire de leau
chaude pour des maisons indi-
viduelles ou des immeubles de
petite taille. Cette gothermie
domestique trs basse nergie
nutilise pas lnergie gother-
mique dorigine interne mais
la chaleur solaire emmagasine
par le sol et le sous-sol.
La gothermie trs basse nergie
devrait se dvelopper dans les
annes venir avec lamliora-
tion des rendements des pompes
chaleur mises sur le march.
Lthiopie se rve en futur gant de la gothermie
Le sous-sol de la valle du Rift recle un potentiel nergtique qui commence seulement
tre exploit.
U
n triple dfi, dmogra-
phique, agricole et nerg-
tique : voil ce qui attend
lthiopie dans les dcennies venir.
Concernant le troisime volet, elle
dispose dune ressource aussi pro-
metteusequemconnue: limmense
potentiel gothermiquequereclent
les entrailles de la valle duRift.
Frapp par la famine dans son ex-
trme sud, le pays, grand comme
deux fois la France et peupl de
88 millions dhabitants, doit aussi
rpondre auxbesoins nergtiques
dune conomie dont la croissance
afcheparadoxalement laplusforte
progression (+ 10 %) du continent
africain, derrire le Ghana (+ 12 %).
Rien nest simple dans la Corne de
lAfrique, surtout que le paramtre
dmographique est loin dy tre
ngligeable : lancien royaume
dAbyssinie devrait compter prs
de150millions dhabitants en2050.
Andersoudresesproblmesdap-
provisionnement ennergie, lthio-
pie sest lance dans la construction
de plusieurs grands barrages, en
dpit parfois de vives tensions avec
les autres pays de la rgion. Lun de
ces projets, baptis Gibe III qui
afche une puissance installe de
1 800 mgawatts (MW) a rcem-
mentfaitlobjetduneptitionsigne
par plus de 400 organisations non
gouvernementales. Celles-ci estiment
quelaralisationdecetouvrage, dans
lesud-ouestdupays, pourraitaffecter
prs de 200 000 personnes vivant
actuellement de lagriculture et de
la pche.
Depuis un demi-sicle, lthiopie
cherche augmenter ses ressources
nergtiques, mais aussi les di-
versier. Elle se tourne aujourdhui
vers les nergies renouvelables. Un
contrat entre le gouvernement et la
socitfranaiseVergnet atsign,
en2009, pour unprojet doliennes
dune capacit de 120 MW dici
2015. Les premires devraient tour-
ner Mkl, danslargionduTigr,
aucours dusecond semestre 2011.
En matire de gothermie, le po-
tentiel quoffre la valle du Rift,
qui stend de la dpression du
Dallol, au nord, au lac Chamo, au
sud, est estim 5 000 MW. La
premire campagne dexploration
gothermique remonte la fin
des annes 1960. Une coopration
entre les autorits de lpoque et le
Programmedes Nations unies pour
le dveloppement (PNUD) a permis
de mener un inventaire des gise-
ments . Seize sites susceptibles
de produire de llectricit grce
dimmenses turbines relies
des rservoirs deau chauffe par
le magma ont alors t recenss.
Le potentiel gothermique est
li lactivit volcanique. Pour
quun site soit exploitable, il faut
que lactivit du magma soit in-
tense , explique SolomonKedebe,
gologue au Geological Survey
of Ethiopia, dpartement de re-
cherche et danalyse dpendant du
ministre des Mines. La valle du
Rift, qui stend de la mer Rouge
la Tanzanie, possde un potentiel
fabuleux. Le Kenya, qui compte
quelques longueurs davance dans
ce domaine, doit dj 11 % de sa ca-
pacit lectrique la gothermie :
environ 170 MW, que le gouverne-
ment entend porter 5 000 MW
dans les cinq prochaines annes.
Le site dAluto Langano, environ
200kmdAddis-Abeba, est exploit
depuis 1999, mais a connu une s-
rie dincidents techniques jusquen
2008. Sa puissance installe nest
actuellement que de 7,3 MW, mais
elle pourrait atteindre 35 MWdici
2015 et 75 MWen 2018.
Les espoirs thiopiens reposent
galement sur le site de Tendaho,
dans la rgion Afar, environ
650 km au nord-est de la capitale.
Cest dans cette zone quasiment
dsertique que se concentrent
la soixantaine de volcans que
compte lthiopie, dont lErta Ale,
le plus actif dentre eux. Un relev
topographique et un forage de re-
connaissance ont montr que la
temprature peut atteindre 300 C
600 m de profondeur. De quoi
situer la puissance potentielle du
site de Tendahoentre 75 et 100MW.
nergie propre capable galement
deproduiredelachaleur oudeleau
chaude, la gothermie est moins
chre que lnergie olienne et
la diffrence de celle-ci disponible
en permanence. Mais la phase ex-
ploratoire ncessite des investisse-
ments levs en comparaison de
lhydrolectricit, par exemplequi
ne peuvent tre assums intgrale-
ment par ltat thiopien.
Legouvernement aconsciencedes
avantages quoffre la gothermie,
une nergie respectueuse de lenvi-
ronnement, assure Hundie Melka
Yadete, chef duGeological Surveyof
Ethiopia. Mais lexploitation de nos
sites dpendaussi de notre capacit
attirer des investisseurs trangers
en matire de recherche et dex-
ploration. Des partenaires islandais,
spcialiss dans la gothermie, se
sont dj montrs trs intresss.
Laduredeviedunsitegothermal
est valueunevingtainedannes.
La salinit du sous-sol ainsi que les
tempratures leves soumettent le
matriel des conditions particuli-
rement rudes. Et lerisquedetrouver
une capacit de production lec-
triqueinfrieureauxprvisionsnest
jamaisexclure. Noussuivonstous
lesprojetsqui favorisent lesnergies
renouvelables, et la gothermie en
fait videmment partie , assure
CheikhDia, charg de missionpour
lAgencefranaisededveloppement
(AFD) Addis-Abeba. Nous avons
dj effectu des visites de secteur
dans le cadre dtudes de faisabilit
et prvoyons aussi des mcanismes
dassurancepour couvrir les risques
dexploitation.
La chaleur des entrailles de la Terre
pourrait termenepas servir seule-
ment la production de llectricit
thiopienne. Les meilleurs sites
gothermiques de la valle du Rift
sont maintenant connus, se flicite
Hundie Melka Yadete. Nous nex-
cluons pas de les utiliser pour diver-
sier nosculturesoudvelopper des
sites touristiques proposant des
cures thermales.
Pierre Lepidi
(21 aot 2011)
POURQUOI
CET ARTICLE ?
Cet articleprsenteunexemple
intressant dexploitation des
ressources gothermiques
danssoncadregodynamique.
En effet, la gothermie peut
constituer une source dnergie
trs intressante pour certains
pays prsentant un fort poten-
tiel gothermique. Cest le cas
delthiopie, qui est situesur la
valle du rift lest de lAfrique.
Cettevalleprsenteuneactivit
volcaniqueintense, lieunere-
montdumagma, dans lecadre
dun contexte godynamique
de rifting continental. Larticle
dresse la liste des diffrentes
sourcesdnergiesditesrenou-
velables que peut exploiter
lthiopie, dont la consomma-
tion nergtique est actuelle-
ment enforteaugmentation. De
par la situation gologique de
l'thiopie, ces projets dusines
gothermiques productrices
dlectricit semblent promet-
teurs. La premire tape, dj
ralise, a consist cartogra-
phier prcisment le potentiel
gothermique de lthiopie. Il
reste maintenant dvelopper
au mieux lexploitation de ces
ressources nergtiques. Ces
projets gothermiques exigent
en effet des investissements
consquents ainsi quedes com-
ptences techniques pointues,
quune coopration internatio-
nale pourrait fournir.
L'ARTICLE DU
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
60
LESSENTIEL DU COURS
61
LESSENTIEL DU COURS
Enjeux plantaires contemporains Enjeux plantaires contemporains
La plante domestique
La domestication lorigine des
plantes cultives
Les espces vgtales actuellement culti-
ves sont issues dun processus de domesti-
cation despces initialement sauvages qui
sest droul sur plusieurs milliers dannes,
comme lattestent les tudes archologiques.
La domestication consiste en une slection arti-
cielle, au cours de laquelle lhomme choisit des
caractres intressants pour la culture et lutilisa-
tion de la plante. Ces caractres sont gnralement
contrls gntiquement et sont donc transmis la
descendance de la plante. Par exemple, les premiers
agriculteurs ont slectionn les grains de bl les
plus lourds pour servir de semences leurs cultures
suivantes. Aprs plusieurs gnrations et slections
successives des grains les plus lourds, les grains pro-
duits ont une masse plus leve que ceux initiaux.
Les caractres des plantes favorables leur utilisa-
tion par lhomme sont en gnral dfavorables la
croissance de la plante en milieu sauvage.
Le croissant fertile (Moyen-Orient), lAmrique
centrale et lAsie du Sud-Est sont les berceaux
de la majorit des espces vgtales cultives
aujourdhui dans le monde entier.
La slection articielle se poursuit
aprs la domestication
Aprs domestication, la slectionarticielle se pour-
suit par diffrentes modalits : slection des carac-
tres intressants, mais aussi hybridations (sponta-
nes ouprovoques) entre deux espces diffrentes,
doublement spontan du nombre de chromosomes
aprs hybridation, mutations spontanes, etc.
Dans le pass, une espce tait souvent soumise
une slection varitale, consistant gnrer diff-
rentes varits de cette mme espce. Chaque varit
prsentait alors des caractres propres, slectionns
enfonctiondes conditions de cultures locales. Lexis-
tence de ces diffrentes varits dune mme espce
vgtalecultiveconstitueuneformede biodiversit.
Actuellement, la cration de nouvelles varits se
L
a culture des plantes pour lalimentation humaine mais
aussi lhabillement, lnergie, la mdecine, etc., constitue
un enjeu majeur pour lhumanit. Au Nolithique ( 12 000
4 000 environ avant J-C), la sdentarisation de lhomme sest
accompagne dune domestication des espces vgtales, qui
sest poursuivie depuis. Comment des plantes sauvages ont-
elles t domestiques par lhomme dans le pass ? Quelles
mthodes lhomme utilise-t-il actuellement pour modier les
plantes quil cultive ?
Mso Amr ique
Mas, haricot, courgette, tomate,
tournesol, cacao
Amr ique du Sud
Tabac, pommedeterre, coton,
manioc, tomate
Avoine, betteravesucrire,
trfle, laitue, lupin, moutarde
Afr ique
Sorgho, melon, bl,
orge
Croissant fer tile
Bl, orge, pois, lin,
oignon, lentille
Asie, Chine
Soja, riz, chou, oignon,
cannesucre, concombre
Asie du Sud-Est, Ocanie
Loriginedespcesvgtalescultivesactuellement
poursuit. Ainsi, dans une espce donne, on peut
confrer un caractre intressant dune varit dite
donneuse une autre varit dite receveuse ,
qui, elle-mme prsente dautres caractres intres-
sants. Pour cela, les deuxvarits sont croises, et les
descendants du croisement prsentant le caractre
dintrt sont slectionns et croiss nouveau avec
la varit receveuse. Aprs rptition des tapes
de croisement et de slection, la varit receveuse
prsentelecaractredintrt, tout enayant conserv
ses autres caractres intressants. Cette amlioration
varitale est un long processus.
Les croisements peuvent aboutir lmergence dune
nouvelle espce. Ainsi la clmentine est-elle un
hybride entre la mandarine et lorange douce.
La biodiversit des plantes cultives
La biodiversit des plantes cultives a, pen-
dant longtemps, t assez importante. Mais
depuis quelques dizaines dannes, la slec-
tion des varits les plus productives aboutit
la culture dun nombre rduit de varits.
Or, cette rduction de la biodiversit peut avoir de
graves consquences. Par exemple, au dbut du
XIX
e
sicle, unchampignon, le mildiou, sattaquaaux
plants de pommes de terre en Irlande et provoqua
une immense famine. la mme poque, lEurope
fut touche par une pidmie duninsecte piqueur,
le phylloxera, qui dcima presque tous les ceps de
vigne. Le vignoble franais fut reconstruit partir de
greffes de cpages franais sur des pieds de vignes
amricains naturellement rsistants au phylloxera.
Il est toujours judicieux, comme le dmontrent
ces exemples, de conserver des varits vgtales
moins productives mais susceptibles dtre por-
teuses de caractres pouvant se rvler intressants,
comme la rsistance des maladies.
Gnie gntique et plantes cultives
Le gnie gntique permet de modier le gnome
dun tre vivant. Les outils du gnie gntique
introduisent un gne dintrt dans le gnome des
cellules dune plante : il sagit dun transfert de
gne ou transgnse. La prsence du gne trans-
fr ou transgne confre la plante de nouveaux
caractres.
La plante ainsi modie est un OGM (organisme
gntiquement modi). Des OGM, comme le mas
transgnique rsistant des herbicides, sont culti-
vs dans plusieurs pays. Les avantages et les incon-
vnients des OGM sont encore largement dbattus
et sont sujets controverse.
Principe du transfert biologique de gnes pour crer des
organismesgntiquement modis(OGM)
Un cal est un amas de cellules vgtales non diffrencies
pouvant gnrer invitrouneplante.
Espce sauvage
nombreuses varits
forte diversit alllique
Premires espces cultivs
Varits cultives
actuelles
Domestication
Slection
varitale
Gnie gntique
(OGM)
Biodiversit
des plantes cultives
Slection par
lHomme des varits
intressantes
+ + +
+
NOTIONS CLS
DOMESTICATION
Processus de slection par
lhomme despces vgtales sau-
vages lorigine des premires
espces cultives.
GNIE GNTIQUE
Ensemble de techniques de mo-
dication du gnome dun tre
vivant sans intervention des
moyens naturels de reproduction.
OGM
Organisme gntiquement modi-
, dont le gnome a t modi
par transgnse, cest--dire par
linsertion de matriel gntique
exogne.
SLECTION ARTIFICIELLE
Choix effectu par lhomme des
espces cultiver en fonction
des caractres jugs intressants
pour lui et qui seront transmis
la descendance de ces espces.
ZOOM SUR...
La rvolution du Nolithique
partir denviron 9 000 avant
J-C, dabord en Orient puis en
Occident, notamment dans le
bassin mditerranen, les popu-
lations connaissent un change-
ment radical de leur mode de vie.
Auparavant chasseurs-cueilleurs,
ils deviennent des producteurs,
pratiquant cultures et levages.
Des crales et lgumineuses
sont slectionnes et cultives,
et des animaux domestiques
sont levs : porc, buf, mou-
ton, chvre, sans oublier le chat,
le chien et le cheval. La chasse
et la cueillette restent toutefois
pratiques.
Cette transformation du
mode de vie, qualifie de r-
volution , a de nombreuses
consquences : amlioration et
diversification de lalimentation,
modification de lhabitat, de la
vie culturelle et spirituelle, etc.
Elle saccompagne galement du
dveloppement de la cramique.
Le Nolithique prend fin avec le
dveloppement de la mtallur-
gie et linvention de lcriture
vers 3 300 avant J-C.
ZOOM SUR...
UN OGM : LE MAS Bt
Lesvgtauxpeuvent treattaqus
par une bactrie appele Agrobac-
teriumtumefasciens. Cettebactrie
porte des petites molcules circu-
lairesdADN, appelesplasmidesTi,
qui sont distinctesduchromosome
bactrien. la faveur dune bles-
sureduvgtal, labactrielinfecte
et injecte ses plasmides Ti dans les
cellules vgtales. Les plasmides Ti
sont alors capables de sintgrer
dans le gnome des cellules de la
plante. Cest ce systme naturel de
transfert de gne qui a t utilis
pour fabriquer les premiers OGM.
Pour la construction de lOGM, le
plasmide Ti est modi. Le gne
responsable de la virulence de la
bactrie est remplac par le gne
codant la toxine dune autre bac-
trie appele Bacillus thurigiensis.
Cette protine est toxique pour
un insecte ravageur, la pyrale. Le
plasmide Ti portant le gne din-
trt (ici celui de la toxine de la
bactrie Bacillus thurigiensis) est
introduit dans des cals. Les cals
sont des amas decellules vgtales
indiffrencies obtenus in vitro.
Seuls les cals ayant intgr le plas-
mide Ti dans leur gnome sont
slectionns et sont alors capables
de gnrer des plantes entires
de mas dont toutes les cellules
sont transgniques. Ainsi, le mas
transgnique obtenu, appel Bt
(car produisant latoxinedeBacillus
thurigiensis), a acquis un nouveau
caractre : capable de synthtiser
une molcule toxique pour la py-
rale, il lui est devenursistant.
UN OGM : LE SOJA
TOLRANT UN HERBICIDE
Plantetransgniquelapluscultive,
cet OGM porte un gne dorigine
bactrienne codant une protine,
lenzyme EPSPS, qui permet la
croissancedelaplanteenprsence
de lherbicide. La plante possde
une autre enzyme EPSPS vgtale
qui est bloque par lherbicide.
Seules les plantes transgniques
ayant lenzymeEPSPSmicrobienne
poussent en prsence de lherbi-
cide, mais pas les autres vgtaux,
commeles mauvaises herbes, ainsi
limins par lherbicide.
DEUXARTICLESDUMONDECONSULTER
Les experts recommandent la prudence
dans le dploiement de certains
OGMp. 63
(Stphane Foucart, 18 novembre 2011)
Ltude qui relance la polmique sur les
OGMp. 64
(Stphane Foucart, 21 septembre 2012)
volutiondelabiodiversitdesespcescultives
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
62
LESSENTIEL DU COURS UN SUJET PAS PAS LES ARTICLES DU
63 Enjeux plantaires contemporains Enjeux plantaires contemporains
Les documents
La carence
1
en vitamine A affecte, daprs lOrga-
nisation mondiale de la sant (OMS), entre 100 et
200millions denfants. Cette carence est responsable
de graves troubles oculaires, de ccit infantile et du
dcs deplus dunmilliondenfants chaqueanne. Or,
les tentatives de diversication nutritionnelle ou de
supplments en vitamines atteignent difcilement
toutes les personnes concernes. Des chercheurs ont
donc travaill sur lenrichissement envitamine A(ou
enprcurseurs devitamineA) decertains aliments de
base dans certains rgimes alimentaires. Ils ont ainsi
mis au point un riz transgnique appel riz dor .
Document 1 : Particularits duriz dor
Le bta-carotne qui, une fois assimil dans le corps
humain, se transforme envitamine Aexiste naturel-
lement dans lenveloppedurizmais pas dans sapartie
comestible, cest--dire lalbumen. Lenveloppe du
riz tant limine de manire amliorer sa conser-
vation, les grains consomms ne contiennent plus
de bta-carotne. Par lintroduction de trois gnes
dans du riz, des chercheurs allemands ont russi
restaurer dans lalbumen une voie de biosynthse
du bta-carotne partir de son prcurseur : le GPP.
Le bta-carotne alors synthtis colore les grains en
jaune, do le surnom de riz dor. Cependant, les
teneurs obtenues jusquprsent ne fourniraient pas
aux populations dmunies en vitamine A les quan-
tits de bta-carotne qui leur seraient ncessaires.
Mais les effets de carences plus ou moins prononcs
pourraient tre sensiblement allgs. Le gnome du
riz dor contient trois gnes codant pour la synthse
denzymes impliques dans la chane de biosynthse
du bta-carotne partir du GPP savoir : deux
gnes de jonquille, qui permettent la fabricationdes
enzymes 1 et 2 ; un gne de bactrie, qui permet la
fabrication de lenzyme 3.
Source : www.maison-des-sciences.ac-versaillers.II.docs/
OGM.pdf
Document 2 : Incertitudes scientiques autour du
riz dor
Le GPP, naturellement prsent
dans le riz, permet la cellule de
fabriquer un certain nombre de
molcules, dont la vitamine E, des
chlorophylles et de lacide gibb-
rellique (substance favorisant la
croissance vgtale). La fraction
du GPP, qui dans le riz dor sera
utilise pour fabriquer du bta-
carotne, ne seraplus disponible pour lasynthse des
autres molcules dont il est galement le prcurseur.
Autrement dit, il est probable que le riz dor, qui fa-
briquedubta-carotne, fabriquemoinsdevitamineE,
et quelesrendementsobtenusavecceriztransgnique
soient nettement diminus en raison dune synthse
amoindrie de chlorophylles et dacide gibbrellique.
Les questions
1.Lebta-carotne,contenunotammentdanslerizdor:
a) permet de pallier les carences enGPP.
b) empche la synthse de la vitamine E de lindividu
qui ingre duriz dor.
c) permetdepallier100%lescarencesenvitaminesA.
d) se transforme en vitamine A chez la personne qui
ingre duriz dor.
2. Lerizdorest issudunetransgnsedetroisgnes
codant pour la synthse :
a) de la vitamine A du riz dans le gnome dune
bactrie.
b) des enzymes permettant la production du bta-
carotne.
c) de la vitamine A dune jonquille dans le gnome
duriz.
d) des enzymes activant la voie de la biosynthse de
la vitamine A.
3.Lafabricationdurizdortransgniqueatfaitepour:
a) pallier les problmes lis lutilisationdepesticides.
b) amliorerlerendementdesriziculturespournourrir
certaines populations humaines.
c) diminuer les effets dune carence alimentaire tou-
chant certaines populations humaines.
e) viter lapport massif dengrais dans les cultures.
4. Daprs certains scientiques, la modication
gntique duriz aboutissant des plants de riz dor
pourrait entraner une production:
a) moindre dufait dunrendement vgtal diminu.
b) accrue de vitamine E par la plante.
c) de plantes plus riches enpigments chlorophylliens.
d) de plantes plus rsistantes auxparasites.
Le corrig
1. d), 2. b), 3. c), 4. a).
Partie 2.1 : Un OGM riche
en prcurseurs de vitamine
GPP
Bta-carotne
Produit
intermdiaireA
Produit
intermdiaireB
Enzyme 1
Enzyme 2
Enzyme 3
La chane de biosynthse
du bta-carotne
Ce quil ne faut pas faire
Cocher deux rponses, ou ne cocher aucune
rponse.
Ngliger les informations apportes par les
documents.
ZOOM SUR...
LORIGINE DES BLS
CULTIVS
Actuellement, nous consommons
du bl tendre, utilis pour les fa-
rines de pain, de pte pizza, et
du bl dur pour la semoule et les
ptes. Quelle est lorigine de ces
deux bls ?
Il y a - 17 000 ans, a eu lieu une
hybridationspontaneentredeux
espces sauvages de crales :
lengrainsauvage (dont le gnome
comporte 2n = 14 chromosomes)
et un gilope inconnu mais
proche de lgilope faux peautre
actuel (comportant galement 2n
=14chromosomes). Cette hybrida-
tionsest accompagne dundou-
blement du nombre de chromo-
somes et a donn naissance une
nouvelle espce : lamidonnier
sauvage, dont les cellules compor-
tent 2n = 28 chromosomes.
Au tout dbut du Nolithique,
vers 12 000 ans, lhomme a
domestiqu cet amidonnier
sauvage. Puis, il a continu de
slectionner des phnotypes de
lamidonnier intressants pour
sa culture. Les caractres de lami-
donnier cultiv ont ainsi t peu
peu modis pour donner, il y
a environ 2 000 ans, le bl dur,
dont le gnome comporte 2n =
28 chromosomes. Dautre part,
vers 9000ans, une hybridation
a eulieuentre lamidonnier culti-
v et lgilope rugueux (espce
2n = 14 chromosomes), suivie
dun doublement du nombre de
chromosomes, qui a abouti la
formationdune nouvelle espce :
lebltendre, dont legnomecom-
porte 2n = 42 chromosomes.
LORIGINE DU MAS
Le mas, originaire du Mexique,
est issu dune domestication, da-
te vers 7 000 ans, dune espce
ancestrale, proche dune espce
sauvage actuelle : latosinte. Suite
cette domestication, les popula-
tions amrindiennes ont procd
une slectionvaritale gnrant
plusieurs varits adaptes lo-
calement. Introduite en Europe
au XIX
e
sicle, le mas est une
des crales les plus produites au
monde, avec le bl et le riz.
Les experts recommandent
la prudence dans le dploiement
de certains OGM
Outre-Atlantique, les avantages lis aux vgtaux tolrants aux herbicides ne sont pas
prennes.
S
ur un sujet aussi sensible
que celui du rapport entre
bnces et risques des bio-
technologies vgtales, les avis
non biaiss par lidologie sont
rares, donc prcieux. Ainsi de
lexpertise rendue publique, mer-
credi 16novembre, sur les effets
agronomiques, environnemen-
taux et socio-conomiques de
la culture des vgtaux tolrants
aux herbicides (VTH).
Ces derniers, dont les plus c-
lbres sont les plants gntique-
ment modis pour rsister au
Roundup lherbicide total de
Monsanto , sont omniprsents
enAmrique. Ils sont rares enEu-
rope, o ils ne sont pas obtenus
par transgnse et ne sont donc
pas considrs comme OGM.
la demande des ministres
chargs de lAgriculture et de
lEnvironnement, cinq scienti-
ques de lInstitut national de la
recherche agronomique (INRA)
et du Centre national de la re-
cherche scientique (CNRS) ont
synthtis lensemble des tra-
vauxscientiques et des donnes
disponibles sur le sujet pour va-
luer le rsultat de ladoption des
VTH, outre-Atlantique, depuis
quinze ans.
Les auteurs se gardent de tout
avis tranch. Mais llment
saillant de leur travail est le
constat que dans les nombreux
pays o les VTH se sont impo-
ss, on na pu que constater
lapparition rapide de formes
adventices (cest--dire de mau-
vaises herbes) rsistantes aux
principales molcules herbicides
quelques annes aprs leur mise
en uvre .
Environ 200 adventices ont
dvelopp ces dernires annes
des rsistances toutes sortes
dherbicides. Unphnomne qui
peut tre ampli par les
conditions dutilisationdes VTH.
Ducoup, les bnces de ces der-
niers, constats court terme,
disparaissent souvent aprs
quelques annes. Les auteurs
constatent que la quantit de
traitements pandre sur des
cultures tolrantes un herbi-
cide total (ou large spectre)
est dabord infrieure celle
ncessaire sur des varits clas-
siques. Les VTH limitent donc
limpact environnemental du
dsherbage.
La libert
des agriculteurs
Mais lapparitiondes rsistances
change la donne. Ce diffrentiel
de consommation dherbicides,
crivent les chercheurs, rgresse
en quelques annes et devient
dfavorable pour le soja et le
coton , rsistants au Roundup.
Inviteaucolloquederestitution,
Marie-Jose Simard, malherbo-
logue lagence canadienne
de recherche agronomique
(Agriculture et Agroalimentaire
Canada) a dress untableauassez
sombre du rsultat de plus de
dix ans dutilisation de cultures
tolrantes au Roundup dans cer-
taines rgions canadiennes.
En Ontario notamment, la
grande herbe poux, Ambrosia
tridis, fortement allergisante et
pourvue dun pollen trs lger,
sest cuirasse face laction de
lherbicide total et pose dsor-
mais dimportants problmes de
dsherbage.
Le bilan de lexprience nord-
amricaine doit-il faire renoncer
une utilisation accrue des VTH
enFrance ? Non, estiment les au-
teurs. Si lanalyse du cas nord-
amricain a mis en vidence des
risques associs une culture
massive de certains VTH, ces r-
sultats ne sont pas directement
transposables la situationfran-
aise , crivent-ils.
La taille des exploitations est
trs suprieure en Amrique du
Nord, explique Michel Beckert
(INRA), coresponsable de lexper-
tise avec Yves Dessaux (CNRS).
Or, cest lune des principales
raisons ladoptionrapide de ces
innovations par les agriculteurs
nord-amricains qui ne peuvent
pas passer beaucoup de temps
sur chaque mtre carr de leur
exploitation.
En dnitive, le principal dfaut
de ces VTH est davoir sduit
trop dagriculteurs. Ils ont t
considrs comme une solution
miracle et cest cela qui se re-
tourne aujourdhui contre eux,
a dclar pour sa part Marion
Guillou, directrice gnrale de
lINRA.
Pour rendre prennes les avan-
tages que prsentent les VTH, il
faut trouver des moyens de rgu-
lationpermettant de limiter leur
utilisation certains crneaux
dusage , estime M. Beckert. Le
d, a conclu Marion Guillou,
sera de mener des recherches
en sciences sociales pour rendre
compatibles ces mcanismes
avec la libert des agricul-
teurs .
Stphane Foucart
(18 novembre 2011)
POURQUOI
CET ARTICLE ?
Cet article a la particularit de
se rapporter la fois au cha-
pitre sur la plante domesti-
que, car il traite des OGM, et
au chapitre sur lvolution et
la slection naturelle. Depuis
plus dune dizaine annes sont
cultivs grande chelle des
OGMprsentant unersistance
un herbicide (ou VTH : vg-
taux tolrants aux herbicides).
Lagriculteur pand lherbicide
auquel est rsistant lOGM,
entranant llimination des
mauvaises herbes ou plantes
adventices. Aujourdhui, un
premier bilan de lutilisation
de ces OGM peut tre dress.
Cet article prsente les re-
commandations dexperts
franais suite ce bilan. Luti-
lisation massive dherbicides
sur plusieurs annes entrane
lapparition de rsistance aux
herbicides parmi les plantes
adventices, ce qui est le rsul-
tat de la pression de slection
exerce par les herbicides sur
ces plantes adventices. Lintrt
dutiliser ces OGM rsistants
aux herbicides diminue donc
avec laugmentation de la r-
sistance aux herbicides. Cette
volution est conforme la s-
lectionnaturelle et tait prdic-
tible. Nanmoins les experts ne
recommandent pas forcment
larrt de la culture des OGM
mais une utilisation plus rai-
sonne de ceux-ci. Rappelons
que, pour linstant, en France,
aucun OGM nest autoris en
agriculture.
1. apport insusant voiremanque
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
64
LES ARTICLES DU
LE MAINTIEN DE LINTGRIT
DE LORGANISME : QUELQUES ASPECTS
DE LA RACTION IMMUNITAIRE
1. La bactrie est
endocyte par le
phagocyte
2. La bactrie est
contenue dans
une vsicule =
le phagosome
3. Le phagosome
fusionne avec un
lysosome riche
en enzymes
4. Digestion de la
bactrie dans le
phagolysosome
5. Exocytose des
lments non
digrs
Plasmocyte
Contamination par
le vir us de la gr ippe
LB
LB
Cytokines
LT
CD4+
Lymphocyte T CD4
auxiliaire activ
LT
auxiliaire
LT
CD4+
LT
auxiliaire
LT
CD8+
Rponse humor ale
Cellule pithliale infecte
BcR
CMH I
TcR
Rponse cellulaire
Cellule
dendr itique
CMH II
Enjeux plantaires contemporains
Ltude qui relance la polmique sur les OGM
Des travaux concluent un lien entre la consommation du mas NK 603 et lapparition
de tumeurs chez le rat.
T
umeurs mammaires chez les
femelles, troubles hpatiques
et rnaux chez les mles,
esprance de vie rduite... Ltude
conduite par le biologiste Gilles-
ric Sralini (universit de Caen)
et parue dans la revue Food and
Chemical Toxicology a dclench
une formidable tempte mdia-
tique. De fait, elle est la premire
suggrer des effets dltres, sur le
rat, de la consommation dun mas
gntiquement modi le NK603
de Monsanto , associ ou non au
Roundup, lherbicide auquel il est
rendu tolrant.
La dmonstration est-elle faite
que cet OGM est toxique ? Voire.
La mme dition de Food and
Chemical Toxicology publie aussi
une tude conduite par Yaxi Zhu
(universit dagriculture de Chine,
Pkin), valuant latoxicitdunmas
rsistant au mme herbicide, sur la
mme souche de rongeurs... mais
sans dtecter de problme.
Loriginalit des travaux de M. Sra-
lini et ses coauteurs est davoir mis
enplaceunprotocoleexprimental
trs ambitieux. Les chercheurs ont
utilis untotal de 200rats pendant
deux ans, contre trois mois en g-
nral pour ce type dexprience. Ils
ont valu les effets dun rgime
alimentaire compos de trois doses
diffrentes du mas transgnique
(11 %, 22 % et 33 %), cultiv ou non
avec son herbicide compagnon.
Trois groupes ont galement t
tests avec des doses croissantes du
produit phytosanitaire seul, non
associ lOGM.
Autotal, donc, ce sont neuf groupes
de vingt rats (trois groupes avec
OGM, trois groupes avec OGM
et Roundup, trois groupes avec
Roundup) qui ont t compars
un groupe tmoin de mme taille,
nourri avec la varit de mas non
transgnique la plus proche de
lOGM test, et sans exposition
lherbicide.
Sur lensemble des groupes traits,
les diffrences les plus signicatives
avec le groupe tmoinapparaissent
au bout denviron un an. Chez les
mles, les congestions et les n-
croses du foie sont 2,5 fois 5,5 fois
plus frquentes. Ces derniers souf-
fraient galement 1,3 fois 2,3 fois
plus datteintes rnales svres. Les
tumeurs mammaires ont t gale-
ment plus frquemment observes
dans les groupes traits mais pas
toujours de manire signicative.
Quant la mortalit, elle a gale-
ment t accrue dans lensemble
des groupes traits. Dans le groupe
tmoin, la dure de vie des mles a
t en moyenne de six cent vingt-
quatre jours, et de sept cents jours
pour les femelles. Aprs que la
priode moyenne de survie sest
coule, toute mort a t largement
considre comme due au vieillis-
sement, crivent les auteurs. Avant
cette priode, 30 % des mles et
20%des femelles dugroupetmoin
sont mortsspontanment, alorsque
jusqu 50 %des mles et 70 %des
femelles sont mortes prmatur-
ment dans des groupes nourris avec
lOGM.
Attention: tous les groupes dani-
maux ne sont compars qu un
groupe tmoin de dix animaux du
mme sexe, explique le biostatisti-
cien Marc Lavielle (Inria). Il aurait
suf quun ou deux rongeurs du
groupetmoinmeurent prmatur-
ment demanirealatoirepour que
la diffrence disparaisse ou ne soit
plus signicative. Statistiquement,
tout cela est trs fragile.
M. Sralini et ses coauteurs notent
que les groupes danimauxexposs
lherbicide seul, de mme qu
lOGM seul, sont affects par les
diverses pathologies releves. Selon
eux, lherbicide pourrait se com-
porter comme un perturbateur du
systmehormonal et produireainsi
des effets non proportionnels la
dose reue.
Quant lOGMseul, satoxicitven-
tuelle pourrait provenir de la modi-
cation dune enzyme (dite ESPS
synthase) produitepar laplanteet
impliquedans lasynthsedacides
amins ayant uneffet deprotection
contre la cancrognse. Le fait que
la synthse de ces acides amins
soit rduite pourrait expliquer, se-
lon les auteurs, les pathologies les
plus frquemment observes chez
les rats tests. Un mcanisme trs
spculatif.
Il faudra que les auteurs rpon-
dent quelques questions qui ne
sont pas dtailles dans leur article,
pointe un spcialiste qui a requis
lanonymat. Comme par exemple
le protocole dapplication du mas
trait, ouencorelhistoriquedelle-
vagedont proviennent les rats, etc.
La souche utilise dite de
Sprague-Dawley, laplus couram-
ment employe dans ce type dex-
prience est en effet connue pour
contracter des cancers mammaires
de manire frquente. Des travaux
publis ennovembre1973dans Can-
cer Research ont ainsi montr que
lincidence de cette pathologie peut
atteindre 45 %chez ces rongeurs, en
labsence de toute intervention.
Je regrette la mdiatisationexces-
sive de ces donnes scientiques,
qui mritent letemps delanalyse ,
estime pour sa part le mdecin et
biochimiste Jean-Christophe Pags,
membre du Haut Conseil des bio-
technologies (HCB) et professeur
luniversit Franois-Rabelais de
Tours.
La prudence est dautant plus de
mise que de nombreuses tudes
de toxicologie ont t menes pour
diffrents OGM et sur diffrentes
espces, sans montrer deffets d-
ltres entre les animaux tmoins
et ceux nourris avec les vgtaux
modis.
Cependant, la plupart de ces tra-
vaux, rassembls dans une analyse
conduite par Chelsea Snell (univer-
sit de Nottingham, Royaume-Uni)
et publie enjanvier dans Food and
Chemical Toxicology, ont t mens
sur des dures infrieures deux
ans et avec un plus faible nombre
de paramtres contrls. Avec des
nancements souvent industriels.
Quant aux quelques tudes de
longue dure dj menes, elles ne
montrent pas non plus deffets bio-
logiques notables, mais ne concer-
nent pas le mas NK603.
Les travaux de M. Sralini dont
le budget est selon lui de plus de
3 millions deuros ont t nan-
cs par la Fondation Charles-
Lopold Mayer, par lassociation
Ceres (qui rassemble notamment
des entreprises de la grande distri-
bution), le ministre franais de la
Recherche et le Criigen (Comit de
recherche et dinformation ind-
pendantes sur le gnie gntique),
association qui milite contre les
biotechnologies.
Stphane Foucart
(21 septembre 2012)
POURQUOI
CET ARTICLE ?
Larticleprsenteltudeportant
sur la toxicit du mas OGM,
effectue par lquipe franaise
du professeur Sralini, parue
enseptembre 2012 et fortement
mdiatise. Conduite pendant
deux annes sur des rats, cette
tude conclut que le mas trans-
gniqueest toxique, entranant la
formationde tumeurs lorigine
dune mortalit accrue de ces
rongeurs. Cette tude est int-
ressante car elle porte sur une
longue priode de temps alors
que les tudes de toxicit des
OGMsont dhabitude limites
6 mois. Mais les instances ayant
expertis ltude de M. Sra-
lini ont conclu que le nombre
danimauxtudis par lot ntait
pas sufsant pour soutenir les
conclusions avances. La repro-
duction de ltude de lquipe
du professeur Sralini dans des
conditions plus rigoureuses
pourrait permettre davancer
dans ce dbat trs anim.
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
66
LESSENTIEL DU COURS
Le maintien de lintgrit de lorganisme : quelques aspects de la raction immunitaire 67
LESSENTIEL DU COURS
Le maintien de lintgrit de lorganisme : quelques aspects de la raction immunitaire
La raction inammatoire,
un exemple de rponse
inne
Les dfenses de lorganisme contre
lintrusion des pathognes
Nous possdons des dfenses mcaniques, comme
limpermabilit de notre piderme et de nos mu-
queuses. Ces dfenses sont compltes par des
dfenses physico-chimiques (larmes, sueur, mucus)
et cologiques (ore bactrienne, prsente natu-
rellement sur et dans notre corps). Malgr tout,
la faveur dune blessure, dune morsure ou dune
piqre, des pathognes peuvent pntrer dans notre
organisme. Une seconde ligne de dfense est alors
oprationnelle pour les neutraliser.
Cette dfense, limmunit inne, se caractrise par
la mise enplace de la ractioninammatoire aigu.
Cette ractioninammatoire aigu est strotype :
elle se droule de la mme faon, quel que soit
lagresseur, et se met en place ds son entre, que
lagresseur soit connu ou non de lorganisme.
La raction inammatoire aigu
Dans les tissus, des cellules comme les mastocytes,
les macrophages, les cellules dendritiques sont
prsentes, en attente dune rencontre ventuelle
avec un pathogne. Ces cellules font partie des
leucocytes (ou globules blancs). Les pathognes
sont dtects par des rcepteurs la surface
de ces leucocytes. La dtection de ces agents
infectieux entrane la scrtion des mdiateurs
chimiques de linflammation par ces leucocytes.
Il existe diffrents mdiateurs chimiques de
linflammation, comme lhistamine, la sroto-
nine, les cytokines pro-inflammatoires (IL-1, IL-6,
TNF-, etc.) et les icosanodes (molcules dont
font partie les prostaglandines, le thromboxane,
la prostacycline et les leucotrines). Ces m-
diateurs chimiques de linflammation activent
la vasodilatation des capillaires.
Les capillaires sanguins se dila-
tent, entranant une augmen-
tation du flux sanguin irriguant
le site inflammatoire. Les cel-
lules des parois des capillaires
scartent, facilitant le passage des
leucocytes du sang vers les tissus, mais aussi de la
lymphe : cela induit chaleur, rougeur et gonfle-
ment. La stimulation des terminaisons nerveuses
voisines provoque une sensation de douleur.
Les leucocytes attirs sur le site inflammatoire
liminent diffremment les pathognes. Les gra-
nulocytes et les monocytes transforms en macro-
phages leur arrive dans les tissus ralisent la
phagocytose du pathogne. La phagocytose est
lingestion et llimination dlments considrs
L
organisme humain est pourvu dun systme de protection
contre les lments pathognes qui menacent son intgri-
t. Ces lments pathognes peuvent tre des virus, des
bactries, des eucaryotes, des parties dtres vivants (pollens,
poils, etc.), des lments physico-chimiques (bres damiante,
poussires, etc.) ou encore les propres cellules modies de
lorganisme, comme les cellules cancreuses. Ces lments pa-
thognes sont potentiellement dangereux pour lorganisme et
le systme immunitaire est charg du maintien de lintgrit de
ce dernier. Comment le systme immunitaire protge-t-il lorga-
nisme des lments pathognes ?
comme trangers par les cellules phagocytaires. La
phagocytose ncessite la reconnaissance du patho-
gne par des rcepteurs membranaires des cellules
phagocytaires (rcepteurs PRR). La membrane du
phagocyte se dforme et emprisonne le pathogne
dans une vsicule, appele phagosome . Cette
vsicule fusionne avec des lysosomes, vsicules
cytoplasmiques contenu acide et riches en en-
zymes. Les enzymes contenues initialement dans
les lysosomes digrent le pathogne. Ainsi la rac-
tion inflammatoire aigu constitue la premire
ligne de dfense active du systme immunitaire
qui vise liminer les pathognes ds leur entre
dans lorganisme.
La raction inflammatoire aigu peut produire des
effets gnants (douleur, gonflement, fivre) voire
dltres pour certains organes si elle se prolonge
trop longtemps. Il peut tre ncessaire dutiliser
des mdicaments qui limitent cette raction
inflammatoire sans limiter le dclenchement
des ractions immunitaires dtruisant les patho-
gnes. Il sagit de mdicaments inflammatoires
comme par exemple l'aspirine ou l'ibuprofne.
Les anti-inflammatoire inhibent la synthse des
mdiateurs chimiques de linflammation. Ces
molcules ont galement une action antalgique,
qui diminue la sensation de douleur et certains
anti-inammatoires font galement baisser la vre.
La rponse inne prpare
la rponse adaptative
Aprs digestion du pathogne, une partie des
molcules restantes sont exposes sous forme de
peptides sur des rcepteurs membranaires des
phagocytes. Ces phagocytes, notamment les cel-
lules dendritiques, quittent le site inflammatoire
et migrent jusquaux ganglions lymphatiques.
Les fragments protiques provenant du pathogne
dtruit, associs aux rcepteurs du phagocyte,
sont alors prsents dautres cellules immuni-
taires, dclenchant ainsi la rponse immunitaire
adaptive, spcifique de lagent infectieux.
Les caractristiques
de limmunit inne
Les mcanismes de limmunit inne, qui existent
chez tous les animaux, sont conservs au cours de
lvolution. Limmunit inne correspond aux
ractions se mettant en place rapidement et se
droulant de manire strotype, quelle que soit
la situation initiale layant dclenche (infection,
lsion des tissus, tumeur, etc.). La raction inam-
matoire aigu est lun des mcanismes essentiel de
limmunit inne. Limmunit inne est fonction-
nelle ds la naissance et ses caractristiques sont
hrites gntiquement. Elle ne ncessite aucun
apprentissage : les ractions de limmunit inne
sont similaires lors de la premire rencontre avec
un pathogne et lors des suivantes. Enn, limmu-
nit inne coopre avec limmunit adaptative, qui
nexiste que chez les seuls vertbrs.
1. La bactrie est
endocyte par le
phagocyte
2. La bactrie est
contenue dans
une vsicule =
le phagosome
3. Le phagosome
fusionne avec un
lysosome riche
en enzymes
4. Digestion de la
bactrie dans le
phagolysosome
5. Exocytose des
lments non
digrs
La phagocytose (daprs Immunologie : le cours deJanis Kuby)
DEUXARTICLESDUMONDE CONSULTER
Trois chercheurs qui ont rvolutionn
la comprhension du systme immuni-
taire. p. 70
(Sandrine Cabut et Herv Morin, 5 octobre 2011)
Laspirine, un nouvel espoir contre le
cancer p. 71
(Paul Benkimoun, 28 mars 2012)
NOTIONS CLS
IMMUNIT INNE
Ensemble des dfenses de lorga-
nismeexistant ds lanaissance, ne
ncessitant pas dapprentissage et
qui semettent enplacetrs rapide-
ment dans des situations diverses.
INFLAMMATION
Ensemble des ractions se pro-
duisant au niveau dune lsion
de lorganisme, o des lments
pathognes peuvent se trouver.
PHAGOCYTOSE
Mcanisme assurant lingestion
par les cellules phagocytaires de
particules ou de cellules lint-
rieur devsicules, les phagosomes,
en vue de leur limination.
REPRE
Les diffrentes cellules
sanguines
Les hmaties, ouglobules rouges,
sont des cellules sans noyau, bi-
concaves et aplaties aucentre. Elles
assurent letransport dedioxygne
dans le sang.
Les leucocytes, ou globules
blancs, interviennent dans la d-
fenseimmunitairedelorganisme:
les granulocytes (galement
appels polynuclaires ) ont
une activit antibactrienne
(diapdse, chimiotactisme et
phagocytose) ;
les cellules dendritiques sont
prsentes dans tous les tissus et en
faible quantit dans le sang. Dans
les tissus, elles dtectent les agents
pathognes et dclenchent larac-
tion immunitaire adaptative ;
les monocytes se diffrencient
en macrophages dans les tissus et
ralisent alors la phagocytose et
la prsentation de lantigne aux
autres cellules immunitaires ;
les lymphocytes participent la
rponse immunitaire adaptative.
Les lymphocytes T sont produits
dans lamoelleosseuseet subissent
leur maturation dans le thymus,
tandis que les lymphocytes B
naissent puis mrissent dans la
moelle osseuse.
Les plaquettes sont des frag-
ments cellulaires qui intervien-
nent dans la coagulation du sang.
REPRE
La dtection dune raction
inammatoire
Une analyse sanguine permet la
dtection de la survenue dune
raction inammatoire chez un
patient. Pour cela, on mesure :
la vitesse de sdimentation (VS)
des hmaties. Linflammation
entrane la production dune pro-
tine, lebrinogne, qui augmente
lagrgationdes plaquettes et donc
la sdimentationdes hmaties. La
vitesse de sdimentationest donc
augmente ;
laprotine-C-ractive(CRP). Cette
protine est produite rapidement
lors de linammation. La mesure
de la concentrationde la protine-
C-ractivepermet destimer quand
est survenue la raction inam-
matoire.
ZOOM SUR
Les maladies inammatoires
chroniques
Ces maladies se caractrisent par
une inammation quasi perma-
nente des tissus touchs. On peut
mentionner :
la polyarthrite rhumatode. Les
articulations sont le sige dune
raction inflammatoire, elles
deviennent douloureuses et d-
formes ;
lamaladiedeCrohn. Elleconsiste
en une inflammation pouvant
toucher la paroi de lensemble
du tube digestif et se manifeste
par des diarrhes et des douleurs
abdominales.
Ces maladies se caractrisent par
des pousses inammatoires trs
douloureuses. La prise en charge
mdicale consiste en des traite-
ments anti-inammatoires visant
rduire les pousses inamma-
toires douloureuses lors des crises.
Des traitements de fond, notam-
ment avec des immunosuppres-
seurs, sont prescrits pour rduire
laractionimmunitaire lorigine
de linammation. Ces maladies
affectent des adultes plutt jeunes
et sont handicapantes dans la vie
quotidienne. Le patient doit tre
suivi toute sa vie : il ny a pas pour
linstant de traitement conduisant
une gurison complte.
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
68
UN SUJET PAS PAS
Le maintien de lintgrit de lorganisme : quelques aspects de la raction immunitaire 69 Le maintien de lintgrit de lorganisme : quelques aspects de la raction immunitaire
Lintitul complet du sujet
Uncamarade sest bless lors dune chute. Quelques
jours plus tard, il a mal, sa plaie est gone, rouge
et purulente ; il consulte un mdecin. Celui-ci,
aprs avoir bien nettoy la plaie, lui donne un
mdicament anti-inammatoire. Votre camarade
ne comprend pas la prescription du mdecin :
Pourquoi dois-je prendre unmdicament puisque
je ne suis pas malade ! , vous dit-il.
Expliquez les mcanismes immunitaires mis en
jeu dans ce cas prcis et lintrt de prendre un
anti-inammatoire. Des schmas explicatifs sont
attendus.
Proposition de corrig
Introduction
Un camarade a fait une chute et sest bless.
Quelques jours aprs quil est tomb, sa plaie
est rouge, gone et du pus sen coule. Son m-
decin a nettoy la plaie et lui a prescrit un anti-
inammatoire. Quels sont les mcanismes de la
raction inammatoire et quel est lintrt de pres-
crire un anti-inammatoire ? Dans une premire
partie, nous allons expliquer notre camarade les
mcanismes de la raction immunitaire, puis, dans
une seconde partie, nous lui expliquerons pourquoi
le mdecin lui a prescrit un anti-inammatoire.
I. Les mcanismes de la raction inammatoire
aigu (ou RIA)
La chute saccompagne de lsions cellulaires avec un
risque lev dentre de micro-organismes patho-
gnes dans lorganisme : une raction immunitaire
aigu se met alors en place trs rapidement.
a. Les symptmes strotyps de la RIA
La ractioninammatoire aigu se caractrise auni-
veaude la plaie par ungonement, une rougeur, une
douleur, unechaleur et uneimpotencefonctionnelle.
b. Le recrutement des dfenses innes lors de laRIA
Des cellules immunitaires, comme par exemple
les mastocytes, les cellules dendritiques ou les
macrophages, sont constamment prsentes aux
lieux dentre potentielle de micro-organismes
dans lorganisme. Par leurs rcepteurs prsents
leur surface, ces cellules immunitaires sont
capables de reconnatre des molcules prsentes
sur la paroi de nombreuses espces de bactries.
Cette reconnaissance entrane la scrtion par ces
cellules immunitaires de molcules : les mdiateurs
chimiques de linammation, comme lhistamine,
la srotonine, les cytokines pro-inammatoires
(IL-1, IL-6, TNF-, etc.) et les icosanodes. Ces mdia-
teurs chimiques de linammationdclenchent une
raction inammatoire aigu. Ainsi lhistamine, en
augmentant la permabilit de la paroi des vais-
seaux sanguins, entrane une vasodilatation et un
afux local de plasma, do la rougeur observe. De
plus, les mdiateurs chimiques de linammation
attirent sur le lieu de linammation les cellules
immunitaires sanguines, comme les granulocytes
et les monocytes sanguins, qui se diffrencient
dans le tissu ls en macrophages.
c. Laction des cellules immunitaires lors de la
RIA
Les cellules immunitaires, comme les granulocytes
et les macrophages prsents dans le tissu ls, ra-
lisent la phagocytose, cest--dire quils ingrent
puis liminent les agents infectieux.
Dans le cas o cette premire rponse immuni-
taire nest pas suffisante pour liminer les micro-
organismes pathognes, interviennent dautres
cellules immunitaires : les cellules dendritiques,
prsentes dans le tissu ls. Ces cellules fixent sur
leurs molcules membranaires du CMH (complexe
majeur dhistocompatibilit) des antignes de
llment infectieux. Ces cellules migrent alors
jusquaux ganglions lymphatiques, o elles pr-
sentent les antignes lis aux molcules du CMH
aux lymphocytes T. Les lymphocytes ainsi activs
interviendront dans l'limination des agents pa-
thognes, dans le cadre de l'immunit adaptative.
II. Lintrt de la prescription dun anti-
inammatoire
Les mdicaments anti-inflammatoires, comme
laspirine, bloquent la scrtion de certains mdia-
teurs chimiques de linammation. Ils permettent
de limiter la vasodilatation, la douleur ou la cha-
leur. Ainsi, les anti-inammatoires permettent
de rduire certains symptmes de la raction
inammatoire aigu sans empcher le droulement
des mcanismes immunitaires qui permettent de
lutter contre les micro-organismes pathognes et
qui sont donc bnques lorganisme.
Partie 1 : La raction
inflammatoire aigu
et lintrt des
anti-inflammatoires
Conclusion
Ainsi, lors dune chute, une raction inammatoire
aigu se met enplace rapidement. Elle se caractrise
par des symptmes strotyps. En faisant interve-
nir des cellules immunitaires et des mdiateurs
chimiques, elle vise liminer les agents pathognes
prsents sur les lsions cellulaires. Si ncessaire, la
RIAdclenche lactivationde limmunit adaptative
qui interviendrapar lasuite. Les anti-inammatoires
permettent de rduire les symptmes de linam-
mation sans diminuer lefcacit de celle-ci. Quels
sont alors les mcanismes immunitaires mis enjeu
dans la rponse immunitaire adaptative ?
Ce quil ne faut pas faire
Ne pas tenir compte des deux parties de la
rponse imposes par lnonc.
Ne pas faire de schma(s).
AUTRES SUJETS POSSIBLES AU BAC
SUR CE THME
tude de documents
tude des mcanismes de la raction inamma-
toire aigu : cellules immunitaires, molcules de
linammation.
tude des modes daction des anti-inamma-
toires.
Mise en vidence de la conservation des mol-
cules de limmunit inne lors de l'volution.
ZOOM SUR...
LES ANTI-
INFLAMMATOIRES
Parmi les anti-inflammatoires,
on distingue :
les corticodes, drivs de
la cortisone. Les corticodes
inhibent la plupart des ph-
nomnes immunitaires, dont
la production des mdiateurs
chimiques de linflammation
comme lhistamine et les ico-
sanodes (molcules drives
de loxydation dacides gras de
la membrane plasmique des
cellules immunitaires, comme
les prostaglandines, le throm-
boxane, la prostacycline et les
leucotrines) ;
les anti-inflammatoires non
strodiens (AINS), comme las-
pirine et libuprofne. Le mode
daction commun des AINS
repose sur linhibition de la
formation dune partie des i-
cosanodes : les prostaglandines.
Les AINS ont donc aussi une
action antalgique (lutte contre
la douleur) et antipyrtique (di-
minution de la temprature de
lorganisme).
noter : le paractamol nest
pas prescrit comme anti-
inflammatoire mais comme an-
talgique et antipyrtique.
LASPIRINE
Depuis lAntiquit, des dcoc-
tions dcorce de saule, conte-
nant de lacide salicylique,
taient utilises contre la
douleur et la fivre. En 1899,
la socit Bayer commercialise
lacide actylsalicylique obtenu
par synthse chimique. Ce nest
quen 1971 quest lucid son m-
canisme daction, qui consiste
en linhibition de la formation
des prostaglandines. En plus de
ses actions anti-inflammatoire,
antalgique et antipyrtique,
laspirine a une action anticoa-
gulante en limitant lagrgation
des plaquettes et peut tre utili-
se pour prvenir des maladies
cardio-vasculaires. Des tudes
rcentes montreraient que las-
pirine faible dose rduirait les
risques de survenue de certains
cancers.
ZOOM SUR...
LES RCEPTEURS
DE LIMMUNIT INNE :
LES PRR
Pendant longtemps, on pensait
que les mcanismes de limmu-
nit ne faisait pas intervenir la
reconnaissance spcifique des
pathognes. Or, dans les annes
1980 furent dcouvertes chez
les cellules de limmunit inne
(mastocytes, macrophages, gra-
nulocytes, cellules dendritiques)
des rcepteurs cellulaires, ap-
pels PRR pour Pattern Reco-
gnition Receptors, capables de
reconnatre les pathognes. Les
lments pathognes prsen-
tent des motifs molculaires
caractristiques appels PAMP
(Pathogen Associated Molecular
Patterns) qui sont reconnus par
les PRR des cellules de limmu-
nit inne.
Les PAMP sont des molcules
abondantes des micro-orga-
nismes : les acides nucliques
(ADN ou ARN) constituent les
PAMP des virus, alors que les
mannanes constituent les PAMP
des champignons. Les PRR re-
connaissant les PAMP sont lo-
caliss la surface des cellules
immunitaires ou lintrieur
de ces cellules : ces localisations
augmentent la possibilit de
rencontre entre les PRR et les
PAMP.
LES RCEPTEURS
DE LIMMUNIT INNE :
DES MOLCULES
TRS CONSERVES
On distingue plusieurs familles
de rcepteurs de limmunit in-
ne ou PRR. Chez la drosophile,
linfection par un champignon
entrane sa reconnaissance par
les rcepteurs appels Toll ,
prsents sur des cellules immuni-
taires de linsecte et dclenchant
llimination de ce pathogne.
Chez les mammifres, une fa-
mille de rcepteurs proches des
rcepteurs de la drosophile a t
identie rcemment : il sagit
des rcepteurs TLR pour Toll Like
Receptors. Ainsi les rcepteurs
de limmunit inne semblent
fortement conservs lors de
lvolution.
UN SUJET PAS PAS
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
70
LES ARTICLES DU
71
LES ARTICLES DU
D
evons-nous tous prendre
de laspirine pour prve-
nir un cancer ? La ques-
tion est de nouveau pose aprs
la publication, mercredi 21 mars,
sur le site de lhebdomadaire m-
dical britannique The Lancet, de
trois nouvelles tudes, menes
par lquipe du professeur Peter
Rothwell. Elles montrent quune
prise quotidienne daspirine r-
duit la mortalit et le risque de
mtastases.
Fondateur de lUnit de recherche
sur la prvention des accidents
vasculaires crbraux luni-
versit dOxford, Peter Rothwell
invite toutefois la prudence :
il est trop tt pour conclure que
tout le monde tirerait un bn-
ce dune telle prophylaxie. Un
avis qui rejoint celui, unanime,
dautres spcialistes.
Les chercheurs britanniques ont
rassembl des donnes issues
dessais destins lorigine
valuer les bnces, sur le plan
cardio-vasculaire, dune prise
en continu daspirine. Cette m-
thode, si elle prsente lavantage
dautoriser facilement laccs aux
donnes de plusieurs dizaines de
milliers dindividus, ne prsente
pas les mmes garanties quune
tude spcifiquement conue
pour tudier les bnces dun
mdicament vis--vis du cancer.
Dans une prcdente tude, pu-
blie en dcembre 2010, M. Ro-
thwell et ses collgues avaient mis
en vidence une rduction den-
viron 20 % du risque de dcs par
cancer chez 25 000 participants
traits quotidiennement pendant
quatre ans avec une faible dose
(75 mg) daspirine. Elle atteignait
30 % 40 % aprs cinq ans de ce
traitement.
Les trois nouvelles tudes conr-
ment la rduction des dcs et
du risque de tumeurs malignes.
Elles montrent aussi une dimi-
nution du risque de cancer avec
mtastases. Les bnfices sont
particulirement vidents sur les
adnocarcinomes, les tumeurs les
plus frquentes.
La premire tude regroupe les
rsultats de 51 essais dans lesquels
les participants taient rpartis, de
manire alatoire, dans ungroupe
avec aspirine ou un groupe sans
aspirine. Le risque de cancer est
diminu de 15 % sous aspirine. La
rduction atteint 37 % avec une
prise pendant cinq ans ou plus. Le
bnce est dj perceptible aprs
trois ans detraitement : lincidence
est alorsrduitedenvironunquart.
Cest norme , reconnat Anne-
Odile Hueber (Institut national de
lasantet delarecherchemdicale,
universit de Nice), spcialiste du
cancer colorectal.
La deuxime tude dmontre
une rduction de 31 % du risque
dadnocarcinome dj mtastas
lors du diagnostic et de 55 % dap-
paritionultrieure de mtastases.
Dans le cas ducancer colorectal, le
risque de mtastases ultrieures
est mme rduit de 74 %.
Laprisequotidiennedaspirineest
aussi associe une diminution
des dcs dus au cancer chez les
personnes qui avaient dvelopp
unadnocarcinome, enparticulier
en labsence de mtastases lors
dudiagnostic (50%de rduction).
Ces rsultats constituent la pre-
mire preuve chez lhomme que
laspirine prvient les mtastases
distancedes cancers , serjouis-
sent les auteurs des trois tudes.
Enfin, la troisime tude situe
la rduction du risque de cancer
colorectal aux alentours de 40%.
Dans uncommentaire qui accom-
pagne les trois tudes du Lancet,
deuxmdecins deHarvardsaluent
ces rsultats incontestables ,
mais soulignent le problme du
risque de saignement inhrent
aux effets de laspirine, qui ui-
die le sang. Un inconvnient qui
tempre les bnces de ce mdi-
cament. LquipedePeter Rothwell
a en effet constat que le risque
hmorragiqueaugmenteaudbut
des essais, mme sil diminue avec
le temps pour disparatre aprs
trois ans de traitement.
Personne ne se hasardera re-
commander sur la seule base de
ces tudes la prise quotidienne
daspirine en prvention du
cancer , affirment en chur
Anne-Odile Hueber, Fabio Calvo,
directeur de la recherche lIns-
titut national du cancer (INCa),
et Dominique Maranichi, ancien
prsident de lINCa.
Pour savoir qui et partir de
quel ge donner de laspirine,
soit en prvention de la surve-
nue dun cancer, soit pour viter
les mtastases ou une rcidive,
il est indispensable de monter
un essai clinique spcique de
grande ampleur. Il appartient aux
organismes publics comme lINCa
et ses homologues dans dautres
pays delemettresur pied, estime
M. Calvo.
Les chercheurs doivent travailler
dans deux directions, selon
Mme Hueber : La premire est
dlucider les mcanismes mol-
culaires expliquant leffet de las-
pirine sur les cellules tumorales
et les tapes initiales du dvelop-
pement duncancer. Lasecondeest
de mettre au point des aspirines
hybrides, qui conserveraient les
avantagesenprventionducancer,
sans les effets indsirables.
Un autre point est claircir.
Nous ne savons pas pourquoi
cela ne marche pas avec une prise
daspirine un jour sur deux , re-
marque M. Calvo. Peter Rothwell
ne serait pas surpris que les ef-
fets dun traitement pris un jour
sur deux, absents court terme,
se rvlent sil est suivi sur le long
terme . Son quipe attend la
publication, lan prochain, des
rsultats dun vaste essai sur las-
pirine en prvention cardio-vas-
culaire, la Women Health Study,
pour trancher cette question.
Paul Benkimoun
(28 mars 2012)
Trois chercheurs qui ont rvolutionn la
comprhension du systme immunitaire
eux trois, ils ont rvolu-
tionn la comprhension
du systme immunitaire,
ouvrant la voie des progrs
dcisifs dans la lutte contre les
infections, les cancers ou encore
les maladies inammatoires. Le
Franais Jules Hoffmann, 70 ans,
lAmricain Bruce Beutler, 53 ans,
et le Canadien Ralph Steinman,
68 ans, sont les trois laurats
2011 du prix Nobel de mdecine,
dcern lundi 3 octobre par le
comit Nobel de Stockholm.
Hoffmann et Beutler, qui se par-
tagent la moiti du prix (dot de
10millionsdecouronnessudoises,
soit 1,08 million deuros), sont r-
compenss pour leurs travaux sur
limmunit inne, premire ligne
de dfense de lorganisme contre
les agressions extrieures. Limmu-
nologisteRalphSteinman, qui avait
obtenulautre moiti duprixpour
sa dcouverte de cellules appeles
dendritiques , impliques dans
limmunit dite spcique , est,
lui, dcd le 30septembre.
Comment les espces vivantes,
et en particulier lhomme,
rsistent-elles aux multiples
attaques, infectieuses et autres,
dont elles font lobjet en perma-
nence ? Les travaux de Jules Hoff-
mann et de Bruce Beutler ont t
dcisifs pour la comprhension
de limmunologie inne , cest-
-dire des tout premiers mca-
nismes de dfense de lorganisme.
Le point commun de ces deux
chercheurs est davoir dcouvert
des rcepteurs qui interviennent
dans la reconnaissance comme
non-soi des germes pathognes
(bactries, champignons) : Jules
Hoffmann chez les insectes, en
1996, et Bruce Beutler chez les
souris, en 1998. Le lien entre
linsecte en loccurrence, la
mouche drosophile , la souris et
lhomme a t rapidement tabli
avec lidenticationde rcepteurs
de type Toll, permettant denclen-
cher la rponse inne chez toutes
ces espces.
tudier la biodiversit
Quant Ralph Steinman, il a d-
couvert, en 1973, des cellules ap-
peles dendritiques , qui inter-
viennent endeuxime ligne. Ces
cellules sont capables de sentir les
dangers extrieurs et de dclen-
cher les rponses immunitaires
dites adaptatives, spcifiques
pour chaque pathogne et qui
nous protgent long terme, pr-
cise SebastianAmigorena, immu-
nologiste et biologiste cellulaire,
directeur de recherche au CNRS.
Les applications de ces travaux
sont normes, dans de multiples
domaines, comme les cancers,
les vaccins synthtiques, les ma-
ladies auto-immunes.
Pour Nicole Le Douarin, pro-
fesseur honoraire au Collge
de France, qui a beaucoup sou-
tenu les recherches de Jules
Hoffmann, ses travaux ont
permis dclairer une question
essentielle reste longtemps
sans rponse, celle de linitia-
tion de la rponse immunitaire,
avant la phase dactivation des
lymphocytes T ou la fabrication
danticorps . Surtout, continue
la chercheuse en embryologie
et biologie, ils ont tabli que
les mcanismes de dfense des
insectes taient conservs chez
des espces plus volues. Cette
persistance dun processus
ancien dans lvolution est un
argument de poids pour tudier
la biodiversit. Il y a une grande
unit dans le monde vivant.
Jean-Luc Dimarcq, qui a pass sa
thse sous la direction de Jules
Hoffmann, salue la vista de
sonancienpatron, qui la conduit
prendre les bonnes dcisions
au bon moment . Rorienter
lensemble des ressources du
laboratoire quil dirigeait vers
ltude de limmunit chez les
insectes tait un pari audacieux.
lpoque, on considrait cette
immunit comme ancestrale,
on hsitait mme employer
ce terme dans les congrs , se
souvient-il. En effet, les insectes
ne disposent pas danticorps,
qui constituaient alors pour la
mdecine le cur du systme
immunitaire. Mais cette ca-
ractristique a sans doute rendu
ltude de limmunit inne plus
facile , note M. Dimarcq.
Strasbourg, entre sablage de
champagne et sollicitations m-
diatiques, Jean-Marc Reichhart,
qui a succd Jules Hoffmann
la tte de lunit 9022 du CNRS,
conrme que cette orientation,
galement porte par sonpouse
Danielle Hoffmann, tait risque.
Pour convaincre nos tutelles,
nous avons eu la chance que
Nicole Le Douarin plaide pour
nous , note-t-il. Il y voit une
morale tirer pour nos ins-
tances : laisser les chercheurs se
lancer dans de telles aventures,
dans des zones quils explorent
sans tre assurs quelles condui-
ront des innovations .
Lquipe de Jules Hoffmann a
peut-tre t en avance sur son
temps en crant, en 1999, une
start-up, Entomed, destine la
production de peptides antimi-
crobiens tirs du vaste arsenal
des insectes. Ces grosses mol-
cules taient difciles synth-
tiser, trop onreuses faire fabri-
quer par des levures : cest l que
le bt a bless , se souvient Ro-
land Lupoli, qui a particip
cette initiative, stoppe en 2005
aprs le retrait dinvestisseurs
amricains. Mais les Hoffman-
niens sont persuads que cette
approche nira par aboutir.
Sandrine Cabut et Herv Morin
(5 octobre 2011)
POURQUOI
CET ARTICLE ?
Cet article dtaille les dcou-
vertes qua rcompenses la
plus haute distinction inter-
nationale dans le domaine de
la mdecine. En 2011, le prix
Nobel de mdecine fut dcern
conjointement trois immu-
nologistes, un Franais, Jules
Hoffmann, un Amricain,
Bruce Beutler et un Canadien,
Ralph Steinman. Jules Hoff-
mann et Bruce Beutler ont fait
progresser les connaissances
sur les mcanismes de limmu-
nit inne. Chez les insectes, o
seule existe la rponse immu-
nitaire inne, Jules Hoffmann
et son quipe ont dcouvert les
rcepteurs Toll capables de re-
connatreles agents pathognes.
Bruce Beutler a identi chez
les mammifres des rcepteurs
TLR (Toll Like Receptors) sem-
blables aux rcepteurs Toll des
insectes. Enn Ralph Steinman
a dcouvert chez lhomme les
cellules dendritiques et leur
importance dans la coopra-
tion entre limmunit inne et
limmunit adaptative. Enfin,
larticlemontrequelarecherche
comporte souvent une part non
ngligeable dinconnu : Jules
Hoffmann choisit de travailler
sur limmunit inne des in-
sectes, sujet qui paraissait bien
peu porteur lpoque et qui se
rvlaloriginedunprixNobel
des annes plus tard.
Laspirine, un nouvel espoir contre le cancer
Le mdicament rduit les risques de mtastases et de dcs, mais sa prescription
gnralise est encore prmature.
POURQUOI
CET ARTICLE ?
Laspirine na pas fini de
nous surprendre ! Utilis
comme anti-inflammatoire
depuis lAntiquit aprs
avoir t extrait du saule,
laspirine a depuis prsent
dautres indications thra-
peutiques. Il est souvent
dit que si laspirine tait
propose actuellement
pour une autorisation de
mise sur le march, celle-ci
lui serait refuse cause
des effets trop divers de
cette molcule. De plus,
larticle propos prsente
les rsultats dtudes met-
tant en vidence le rle
de protection de laspirine
contre les cancers. Ces r-
sultats trs prometteurs ne
sont nanmoins que prli-
minaires. Dautres tudes
doivent tre menes avant
de formuler des recomman-
dations thrapeutiques,
tant donn le risque
hmorragique important
qui accompagne la prise
prolonge daspirine. La
dcouverte de lorigine de
la protection contre le can-
cer que confre laspirine
pourrait galement ouvrir
la voie un nouveau champ
thrapeutique dans la pr-
vention des cancers.
Le maintien de lintgrit de lorganisme : quelques aspects de la raction immunitaire Le maintien de lintgrit de lorganisme : quelques aspects de la raction immunitaire
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
72
LESSENTIEL DU COURS
Le maintien de lintgrit de lorganisme : quelques aspects de la raction immunitaire 73
LESSENTIEL DU COURS
Le maintient de lintgrit de lorganisme : quelques aspects de la raction immunitaire
Limmunit adaptative,
prolongement de
limmunit inne
La rponse immunitaire adaptative
Suite la raction inammatoire, des cellules den-
dritiques capturent sur les sites de linfectionlagent
infectieux, dont les fragments protiques consti-
tuent des antignes. Sur ces cellules dendritiques,
les antignes sont exposs au sein de molcules
spciques appeles complexe majeur dhisto-
compatibilit (CMH). Les cellules dendritiques
migrent vers des ganglions lymphatiques, o elles
rencontrent de nombreux lymphocytes T (LT).
Les LT expriment leur surface des rcepteurs T,
appels TCR (TCell Receptors), associs dautres
molcules de surface comme CD4 ou CD8. Les
LT CD4+
portent le rcepteur CD4+
et le TCR, qui est
identique pour un mme clone de LT CD4+
. Le TCR
des LT CD4+
interagit avec le peptide antignique
prsent par le CMHdes cellules dendritiques. Cette
reconnaissance spcique entrane lactivation des
LT CD4+
, qui scrtent alors des cytokines, comme
linterleukine 2 (IL2). IL2 est un mdiateur soluble
de communicationqui induit la prolifrationclonale
puis la diffrenciation des LT CD4+
layant secrte
en lymphocytes T auxiliaires (LT helpers ou LTh).
Ainsi, les cellules dendritiques sont des cellules
prsentatrices de lantigne activant la formation
de LT auxiliaires, dont la fonctionest de scrter des
molcules activant les cellules immunitaires.
Les lymphocytes B (LB) portent leur surface un
rcepteur appel BCR (B Cell Receptor), qui
correspond un anticorps membranaire. Tous les
anticorps membranaires dun mme LB ou dun
mme clone de LB sont identiques.
La reconnaissance spcique entre un BCR et lanti-
gne correspondant entrane lactivation du LB. Les
LT auxiliaires, prsentant la mme spcicit pour
lantigne que les LB, scrtent linterleukine 2, qui
stimule la prolifration clonale puis la diffrencia-
tion de ces LB en plasmocytes scrteurs danti-
corps. Les anticorps scrts par les plasmocytes
issus dun mme clone de LB ont tous la mme
spcicit pour lantigne.
Un anticorps ne reconnat quun seul type dan-
tigne. Lanticorps possde, en effet, une partie
constante et une partie variable, qui reconnat
lantigne. Les anticorps permettent la neutrali-
sation de lantigne. Ils sont produits en grande
quantit par les plasmocytes et se xent sur les virus.
Cette xation empche les virus dinfecter leurs
cellules-cibles. La partie constante des anticorps
est reconnue par des rcepteurs membranaires
des cellules phagocytaires, qui liminent lagent
infectieux recouvert danticorps par phagocytose.
Ainsi, lintervention des LB scrteurs danticorps
ou plasmocytes constitue limmunit adaptative
mdiation humorale, spcique
de lantigne et qui ncessite lin-
tervention des LT auxiliaires.
Les cellules infectes par des virus,
sont dtruites par les lympho-
cytes T cytotoxiques (LTc). Dans
les ganglions lymphatiques, des
cellules prsentatrices de lanti-
gne expriment au sein de leur
CMH lantigne, qui est reconnu
spcifiquement par le TCR des
LT, porteurs galement du rcep-
teur CD8. Cette reconnaissance
entrane lactivation des LT CD8
+
,
qui prolifrent puis se diffren-
L
immunit inne est prsente chez tous les tres vivants.
Mais chez les vertbrs sest dveloppe une rponse im-
munitaire plus spciquement dirige contre le pathogne
et dote dune mmoire : limmunit adaptative. Cette rponse
se met en place en parallle de la rponse inne, avec quelques
jours de dlai. Quelles en sont les caractristiques ?
Chronologie de la rponse immunitaire
cient en LT cytotoxiques. Cette prolifration et
cette diffrenciation ncessitent la stimulation par
linterleukine 2, secrte par les LT auxiliaires ac-
tivs par le mme antigne. Les LT cytotoxiques se
xent par leur TCR lantigne prsent au sein du
CMH de la cellule infecte et librent des molcules
cytotoxiques la dtruisant. Les LT cytotoxiques
constituent limmunit adaptative mdiation
cellulaire, spcique de lantigne et ncessitant
les LT auxiliaires.
Le VIH induit un syndrome dimmunodcience
acquise (sida)
Le virus de limmunodcience humaine (VIH)
infecte et dtruit diffrentes cellules, dont princi-
palement les LT CD4+
. Le patient est dit sropositif
pour le VIHquand il produit des anticorps anti-VIH.
Pendant quelques annes, le systme immunitaire
fait diminuer le taux de virus dans le sang. Mais le
VIH, toujours prsent dans lorganisme, dtruit les LT
CD4+
, dsorganisant le systme immunitaire. Le pa-
tient est dit atteint dusidaquandil dveloppe des
maladies opportunistes, pouvant entraner sondcs.
Les trithrapies actuelles retardent lvolution de la
maladie mais nradiquent pas le VIHde lorganisme.
La maturation du systme
immunitaire
Les LB et LT sont produits dans les organes lym-
phodes primaires (thymus, moelle osseuse). La
grande diversit des rcepteurs BCR des LB et TCR
des LT, capable de reconnatre une multitude danti-
gnes, est le rsultat alatoire
de mcanismes gntiques
complexes. Puis, les LB et LT
capables de reconnatre les
constituants de lorganisme et
dits auto-ractifs, sont limi-
ns. lissue de cette slection
ngative, les LB et LT sont dits
nafs : ils nont pas encore ren-
contr leur antigne. Ces lym-
phocytes migrent dans les or-
ganes lymphodes secondaires
(comme les ganglions lympha-
tiques), oils sont susceptibles
de rencontrer leur antigne. La
maturation du systme im-
munitaire est donc le produit
dun quilibre dynamique
entre production de cellules
immunitaires et limination
des cellules immunitaires auto-
ractives. La maturation du
systmeimmunitaireest lvo-
lutiondurpertoire immuni-
taire au cours de la vie de lin-
dividu, en fonction des
antignes rencontr, et consti-
tue un apprentissage.
Structure dunanticorps
Plasmocyte
Contamination par
le vir us de la gr ippe
LB
LB
Cytokines
(IL2)
Lymphocyte T auxiliaire
activ
LT
auxiliaire
LT
CD4+
LT
LT
cytotoxique
Rponse humor ale
Cellule pithliale infecte
CMH
TCR
Rponse cellulaire
Cellule
dendr itique
CMH
LT
auxiliaire
BCR
Secr tion
danticor ps
Neutr alisation
du vir us
Mor t des cellules infectes
CD8+
Lutte contre le virus de la grippe
MOTS CLS
ANTICORPS
Glycoprotines synthtises
par les plasmocytes que lon
retrouve dans le plasma et dans
dautres liquides biologiques.
Les anticorps sont aussi appels
immunoglobulines car ils
sont retrouvs aprs lectro-
phorse dans des protines du
srum, dans les diffrentes frac-
tions (, et ) des globulines.
Chez lhomme, les anticorps
ont pour forme un Y compos
de quatre chanes peptidiques
(deux chanes lourdes iden-
tiques et deux chanes lgres
identiques). Chaque anticorps
possde deux sites de fixation
aux antignes situs aux ex-
trmits variables des chanes
lgres et des chanes lourdes.
On distingue cinq classes dim-
munoglobulines : IgG, IgM, IgA,
IgE et IgD, en fonction de la na-
ture de la rgion constante de la
chane lourde. La sropositivit
pour un antigne donn cor-
respond la prsence dans le
srum danticorps dirigs contre
cet antigne et signe l'activa-
tion du systme immunitaire
en rponse la prsence d'un
antigne.
ANTIGNE
Toute structure molculaire
pouvant tre reconnue par un
rcepteur de limmunit adap-
tative, soit un anticorps, soit
un TCR (rcepteur des LT), soit
un BCR (rcepteur des LB) et
dclenchant ainsi une rponse
immunitaire adaptative.
CELLULES
PRSENTATRICES
DANTIGNE (CPAg)
Les CPAg constituent une cat-
gorie de phagocytes capables
dexprimer leur surface des
dterminants antigniques lis
des protines du complexe
majeur dhistocompatibilit
(CMH). Il sagit, par exemple,
des cellules dendritiques et des
macrophages. Ces cellules sont
au cur de la coopration entre
limmunit inne et limmunit
adaptative.
CHIFFRES
CLS
Le sida en France
Au total, en 2009, 83 000 per-
sonnes ont dvelopp un sida de-
puis le dbut de lpidmie dans
les annes 1980, et 46 000 ma-
lades en sont dcds. En 2009,
1 450 nouveaux cas de sida ont
t dclars.
En 2009, on estime environ
152 000 le nombre total de per-
sonnes infectes en France par
le VIH, dont environ 50 000 ne
connatraient pas leur sropo-
sitivit.
Environ 6 300 personnes ont
dcouvert leur sropositivit
en 2010 : ce nombre est stable
depuis quelques annes. 30 %
des diagnostics de sropositi-
vit pour le VIH sont tablis
un stade tardif, alors que le taux
de LT CD4+
est dj faible. Le
diagnostic prcoce ne concerne
que 36 % des sropositivits d-
tectes.
ZOOM SUR
La transmission du virus
du sida et la prvention
La transmission du sida a lieu :
lors de rapports sexuels (80 %
des cas). Prvention par utilisa-
tion de prservatifs masculin ou
fminin ;
lors dune contamination par le
sang (transfusion sanguine, se-
ringue contamine). Prvention
par utilisation de matriel strile
et usage unique ;
de la femme enceinte lenfant,
lors de la grossesse, laccouche-
ment ou lors de lallaitement.
Dans les pays industrialiss, les
traitements ont rduit presque
totalement le risque de la trans-
mission de la mre lenfant.
Prvention par prise en charge
mdicale pendant la grossesse,
naissance par csarienne, utilisa-
tion exclusive de lait articiel et
traitement post-natal de lenfant.
De fausses croyances persistent
sur la transmission du virus. Il
ne peut pas se transmettre par
la salive, la sueur, les larmes, les
piqres dinsectes ou encore le
toucher.
DEUXARTICLESDUMONDE CONSULTER
Limmunothrapie, une nouvelle arme
contre les cancers p. 76-77
(Florence Rosier, 8 novembre 2012)
Une piste pour radiquer le VIH de lorga-
nisme p. 77
(Paul Benkimoun, 17 mars 2012)
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
74
UN SUJET PAS PAS
Le maintien de lintgrit de lorganisme : quelques aspects de la raction immunitaire 75 Le maintien de lintgrit de lorganisme : quelques aspects de la raction immunitaire
Lintitul complet du sujet
On sait que les greffes de tissus ne sont possibles
que si le donneur et le receveur sont compatibles.
Oncherche prciser les mcanismes immunitaires
impliqus dans le rejet dune greffe de peau chez
la souris.
Exploitez lensemble des rsultats exprimentaux
proposs dans le document et montrez quils sont
en accord avec lhypothse selon laquelle le rejet
de greffe chez la souris repose sur des mcanismes
dimmunit adaptative impliquant des effecteurs
cellulaires.
Le document
Quelques rsultats exprimentauxchez les souris
Des greffes de peauont t ralises chez des souris
de lignes pures (homozygotes pour tous leurs
gnes) appeles lignes A et lignes B. On observe
que :
ungreffonde peauissudune souris de ligne Bim-
plant une souris de ligne Best toujours accept ;
un greffon de peau issu dune souris de ligne A
implant une souris de ligne B est parfaitement
fonctionnel sixjours aprs la greffe, mais totalement
dtruit au bout de onze jours ;
une souris de ligne Bayant prcdemment rejet
un premier greffon issu dune souris de ligne A
rejette un deuxime greffon de souris de ligne A
en six jours.
Des souris de ligne Bsont diteshyperimmunises
lorsquon leur a greff trois reprises, trois se-
maines dintervalle, de la peaude souris de ligne A.
Les chercheurs prlvent alors chez ces souris dune
part leur srum (plasma sanguin) et dautre part
Partie 2.1 : Greffe de
peau chez la souris
des cellules lymphodes dans les ganglions lympha-
tiques situs prs du greffon. Des souris de ligne B
sont dites neuves (notes BN) si elles nont subi
aucun traitement.
Exprience 1
Des souris de ligne B neuves (BN) reoivent le
srumdes souris de ligne Bhyperimmunises, puis
trois jours plus tard une greffe de peau de souris de
ligne A. Onze jours plus tard, le greffon est rejet,
alors quil tait entirement fonctionnel jusquau
sixime jour.
Exprience 2
Dautres souris BNreoivent des injections aujour 1,
une greffe de peau issue dune souris de ligne A au
jour 3. Ltat du greffon est observ au jour 6. Les
rsultats sont les suivants :
Proposition de corrig
Une greffe de peauentre une souris donneuse et une
souris receveuse appartenant une mme ligne
homozygote nest jamais rejete : les deux souris
sont donc compatibles.
Une greffe de peau dune souris donneuse de la
ligne A une souris receveuse de la ligne B est
fonctionnelle au sixime jour mais est rejete au
onzime jour : les souris ne sont pas compatibles.
La greffe de peau dune souris donneuse de la
ligne A une souris receveuse de la ligne B qui a
dj rejet un premier greffon issu de A est rejete
beaucoup plus rapidement, en six jours.
Exprience 1
Des souris de ligne B neuves (souris NB) nayant
reu aucun traitement au pralable reoivent le
srum des souris de ligne B hyperimmunises,
puis reoivent trois jours plus tard une greffe de
peau de souris de ligne A : la greffe nest rejete
quau onzime jour. Les molcules contenues dans
le srum des souris de ligne B hyperimmunises
ne permettent pas un rejet prcoce de la greffe au
sixime jour chez les souris NB, comme il est observ
chez les souris de ligne B hyperimmunises.
Exprience 2
Des souris BN reoivent les cellules lymphodes vi-
vantes des souris de ligne Bhyperimmunises, puis
reoivent trois jours plus tard une greffe de peaude
souris de ligne A: le greffonest rejet ds le sixime
jour. Les souris BNayant reules cellules lymphodes
vivantes des souris de ligne B hyperimmunises
ragissent la greffe comme les souris de ligne B
hyperimmunises, comme si elles avaient dj t
en contact avec le greffon et l'avaient rejet.
De plus, la greffe des souris BNayant reules cellules
lymphodes tues des souris de ligne B hyperim-
munises entrane un greffon encore fonctionnel
au sixime jour. Les cellules lymphodes injectes
doivent donc tre vivantes pour entraner le rejet
du greffon au sixime jour.
Enn, la greffe des souris BN ayant reu les cellules
lymphodes vivantes des souris de ligne B non
immunises entrane ungreffonencore fonctionnel
au sixime jour. Pour entraner un rejet prcoce du
greffon, les cellules lymphodes injectes vivantes
doivent provenir de souris hyperimmunises.
Ainsi, les cellules lymphodes contenues dans les
ganglions lymphatiques des souris sont respon-
sables du phnomne de rejet lors des greffes entre
donneur et receveur non compatibles. La rponse
immunitaire est dautant plus rapide que le receveur
a dj t en contact avec lantigne : cela justie
donc le caractre adaptatif de cette rponse immu-
nitaire.
Ce quil ne faut pas faire
Se contenter de saisir les donnes du document
sans montrer comment les rsultats exprimen-
taux permettent de valider lhypothse formule
par lnonc.
AUTRES SUJETS POSSIBLES AU BAC
SUR CE THME
Matrise des connaissances :
questionde synthse
Prsenter les ractions immunitaires de limmu-
nit adaptive en s'appuyant sur lexemple dune in-
fectionvirale, la grippe.
Prsenter le recrutement des LB et leur fonction
dans lliminationdes agents pathognes.
Prsenter le recrutement des LT cytotoxiques et
leur fonction dans llimination des cellules infec-
tes par des virus.
En s'appuyant sur lexemple du sida, montrer le
rle central des LT auxiliaires dans la rponse im-
munitaire adaptative.
ZOOM SUR
LE COMPLEXE MAJEUR
DHISTOCOMPATIBILIT
Le complexe majeur dhistocom-
patibilit (CMH) est lensemble
de six gnes principaux (A, B, C,
DR, DQ, DP) situs, chez lhomme,
sur la paire de chromosomes 6
et codant des glycoprotines
de surface qui interviennent
dans le rejet de greffes et la
reconnaissance du soi modi.
Chez lhomme, le complexe
majeur dhistocompatibilit est
appel systme HLA (Hu-
man Leucocytes Antigens). Le
complexe majeur dhistocom-
patibilit forme un ensemble de
gnes trs polymorphes de par
un nombre lev dallles : le
complexe majeur dhistocom-
patibilit nest jamais identique
chez deux individus (sauf chez
les vrais jumeaux).
LA GRANDE DIVERSIT
DES RCEPTEURS
DES CELLULES
IMMUNITAIRES
Limmunit adaptative gnre,
tout au long de la vie, un trs
grand nombre de rcepteurs BCR
des lymphocytes B et TCR des
lymphocytes T. Limmensit des
rpertoires B et T permet poten-
tiellement de faire face une
multitude dantignes diffrents.
La spcicit des lymphocytes B
et lymphocytes T, fonde sur celle
des rcepteurs quils expriment,
est obtenue grce unrarrange-
ment au hasard de linformation
gntique : au cours de la ma-
turation des lymphocytes dans
la moelle osseuse (pour les LB)
et dans le thymus (pour les LT),
les gnes codant les rcepteurs
sont construits par combinai-
son alatoire de segments de
gnes prsents dans les futurs
lymphocytes B et lymphocytes T.
Au cours de ce processus, les
lymphocytes perdent une partie
de leur gnome. Les possibilits
de ces rarrangements sont esti-
mes 10
18
, alors que seulement
10
12
10
15
lymphocytes sont pr-
sents ou produits dans le corps.
Ce rarrangement des gnes
samorce avant la naissance.
ZOOM SUR
LA GREFFE DORGANES
La greffe ou transplantation
dorganes est le remplacement
dun organe ou dun tissu dfec-
tueux par un organe ou un tissu
sain, appel greffon et prove-
nant de la personne greffe ou
dun autre donneur. La greffe
dorganes est parfois la seule
issue thrapeutique lorsque les
organes vitaux sont atteints de
manire irrversible.
En 2011, en France, 40 000 per-
sonnes vivent avec un greffon
fonctionnel. 4 945 personnes ont
t greffes en 2011 : le rein est
la greffe la plus courante (60 %),
suivie de celle du foie (26 %), du
cur (8 %), puis du poumon et
du pancras. De 1991 2011, le
nombre de greffes a augment
de 41 % mais cette augmentation
ne suffit pas rsorber lattente :
16 371 patients taient en at-
tente dun greffon en 2011, soit
presque le double par rapport
1997.
Le greffon tranger lorganisme
peut dclencher une raction
immunitaire de rejet. Le rejet
peut tre aigu, ou chronique, en-
tranant, terme, la destruction
du greffon ou une perte de sa
fonctionnalit. Les traitements
immunosuppresseurs ont pro-
gress ces dernires annes, mais
ils s'accompagnent d'un risque
accru dinfections et de tumeurs
chez le patient. Les recherches
se poursuivent pour augmenter
la tolrance du systme immu-
nitaire pour le greffon, tout en
conservant une ractivit vis-
-vis des autres antignes.
LES MALADIES
AUTO-IMMUNES
Les maladies auto-immunes sont
dues lattaque et la destruc-
tion des cellules de lindividu
par son propre systme immu-
nitaire. Par exemple, le diabte
juvnile ou insulino-dpendant
est d lattaque des cellules du
pancras responsables de la syn-
thse dinsuline ; la sclrose en
plaques correspond lattaque
de la gaine de myline entourant
les cellules nerveuses.
UN SUJET PAS PAS
Infection au jour 1 Rsultat au jour 6
Greffe
au jour 3
de cellules lymphodes vivantes de souris
de ligne B hyperimmunises
de cellules lymphodes tues de souris de
de ligne B hyperimmunises
de cellules lymphodes vivantes de souris
de ligne B non immunises
peau de souris
de ligne A
peau de souris
de ligne A
peau de souris
de ligne A
la majorit des greffons sont dtruits ou
prsentent des ncroses partielles
les greffons sont toujours fonctionnels
les greffons sont toujours fonctionnels
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
76
LES ARTICLES DU
77
LES ARTICLES DU
Le maintien de lintgrit de lorganisme : quelques aspects de la raction immunitaire Le maintien de lintgrit de lorganisme : quelques aspects de la raction immunitaire
D
ans la guerre sans merci
dploye contre les can-
cers, une multitude dof-
fensives mobilisent nos lignes de
dfense immunitaire. Ce sont les
immunothrapies des cancers .
Les premires armes de ce combat
ont t fourbies en 1891 par un
chirurgien new-yorkais : Chez de
jeunes malades atteints dosto-
sarcome, une tumeur des os trs
agressive, William Coley constate
quune infectionaigu provoque la
rgression des tumeurs , raconte
Laurence Zitvogel, oncologue m-
dicale lInstitut Gustave-Roussy
(IGR) de Villejuif (Val-de-Marne).
En 1893, il injecte ces malades
un cocktail de bactries inactives
qui font leur tour rgresser ces
cancers. Comment ? En stimulant
le systme immunitaire, qui li-
bre son agressivit vis--vis des
tumeurs.
tonnamment, cette approche
constitue encore le traitement
standard de certains cancers de la
vessie depuis quarante ans : aprs
rsectionchirurgicale de la tumeur,
des injections rptes de bacille du
BCGsont pratiques dans la vessie.
Chez les patients, plusieurs observa-
tions conrment le rle primordial
du systme immunitaire dans le
contrle des cancers oumontrent
ses dfaillances dans leur dvelop-
pement. Cest dabord le constat
dune frquence accrue de certains
lymphomes chez les patients
immunodprims ( limmunit
affaiblie par une maladie ou un
traitement).
Cest ensuite cette dcouverte dans
des cancers du clon, du sein ou
de lovaire : Plus le site primitif
de la tumeur est riche en certaines
cellules immunitaires, moins le pa-
tient fait dercidiveet plus sasurvie
est longue , explique FranckPags,
de lhpital europen Georges-
Pompidou (Paris). Avec lquipe
Inserm de Jrme Galon, il a mis
au point un score immunitaire
vise pronostique, fond sur le
nombre de lymphocytes qui inl-
trent les tumeurs colorectales.
terme, ce score pourrait permettre
dadapter les traitements.
Mais mesure que la tumeur crot,
elle sallie rapidement un rgi-
ment transfuge de limmunit : les
cellules T dites rgulatrices , qui
freinent loffensive destructrice
des cellules T cytotoxiques. Ces
manuvres flonnes sappuient
sur plusieurs molcules cls, telle
la protine CTLA4. Lorsque celle-ci
apparat ennombre la surface des
cellules T, elle retarde oufreine lac-
tion des cellules cytotoxiquesqui
ne peuvent poursuivre leur attaque
tumorale.
Ltape suivante a t le dve-
loppement de stratgies visant
contrecarrer cetteimmunosuppres-
sion, relateLaurenceZitvogel. Do
la mise au point dune premire
molcule thrapeutique : un an-
ticorps anti-CTLA4, lipilimumab.
Chef de le des immunothrapies
des cancers, il est aujourdhui
approuv dans le traitement des
mlanomes mtastatiques par les
agences dumdicament amricaine
et europenne. Avec lipilimumab,
la survie des patients est au moins
double. Chez les rpondeurs, cette
survie apparat durable, avec un
recul de quatre cinqans, se rjouit
Cleste Lebb*, onco-dermatologue
lhpital Saint-Louis, Paris. Lim-
munothrapie a longtemps t un
serpent demerdans lemlanome,
mais aprs quinze ans de dcep-
tions a marche enn !
Pour autant, ce nest pas la pana-
ce. Le cot initial demand par le
laboratoire (Bristol-Myers Squibb)
tait si lev 80 000 euros par
patient que les ngociations avec
les autorits sanitaires bloquent.
Pour lheure, lipilimumab nest
pas rembours. Certains hpitaux
franais acceptent de le nancer,
mais beaucoup refusent.
Autres faiblesses de lipilimumab :
Nous ne disposons pas encore de
biomarqueurs pour prdireles 20%
depatients qui rpondront, regrette
Caroline Robert*, onco-dermatolo-
gue lIGR. Et lipilimumab a un
prol deffets indsirables trs inha-
bituel. Parce quil libre le systme
immunitaire, il peut dclencher une
auto-immunit en sattaquant aux
tissus sains. Il faut bien connatre
ces effets, en informer les patients
et apprendre les grer.
Dautres immunothrapies sont en
cours dvaluation. Parmi elles, un
anticorps anti-PD1, qui rveille
le systme immunitaire en dblo-
quant undeses verrous. Aucongrs
mondial de cancrologie qui se te-
nait dbut juin Chicago, lASCO,
cet anticorps a t lu molcule
de lanne . Il a donn lieu deux
articles et un ditorial, publis
en juin dans la prestigieuse revue
NewEnglandJournal of Medicine. Des
essais prliminaires suggrent des
taux de rponses prolonges chez
20%30%des patients dans des tu-
meurs trs agressives : mlanome,
cancer dureinet cancer dupoumon
non petites cellules.
Une autre stratgie est dduquer
les lymphocytes T nafs du patient
avec les antignes isols de son
propre cancer , explique Laurence
Zitvogel. Chez des patientes at-
teintes de cancer du col de lutrus,
cette vaccinationthrapeutique
donne des rsultats prometteurs.
Les antignes administrs sont
issus des papillomavirus de lher-
ps HPV16 et 18, en cause dans le
dveloppement de ces cancers. Et
dans un article publi, n 2009,
dans le New England, lquipe de
Cornelis Melief tablit lefcacit de
la vaccination thrapeutique (par
des longs peptides de ces virus)
dans des cancers de la vulve. Le
11 octobre, uneautrequipemontre,
dans Science Translational Medicine,
que des femmes atteintes de l-
sions prcancreuses dclenchent
une rponse immune anticancer
spcique et durable aprs avoir
reu, par lectroporation, des frag-
ments dADN (des oncognes des
virus HPV16 et 18). Plusieurs autres
vaccins thrapeutiques sont lessai
contre des mlanomes, des cancers
de lovaire ou du sein.
Les chercheurs mobilisent aussi
Limmunothrapie, une nouvelle arme contre les cancers
Plusieurs observations conrment le rle primordial du systme immunitaire dans le
contrle de ces maladies. Diffrentes thrapies sont dj disponibles, dautres sont lessai.
Une piste pour radiquer le VIH de lorganisme
De nouveaux traitements pourraient liminer les rservoirs viraux dans lesquels le
virus du sida persiste.
I
l est jusquici impossible,
malgr un traitement an-
tirtroviral, dradiquer de
lorganisme le virus de limmu-
nodcience humaine (VIH). Cet
objectif deviendra-t-il ralisable
une chance de cinq dix ans ?
Lventualit a t voque lors de
la Confrence sur les rtrovirus
et les infections opportunistes
(CROI) qui sest tenue Seattle
(tats-Unis) du 5 au 8 mars. Le
professeur Christine Rouzioux,
virologue lhpital Necker (Pa-
ris), a prsent une synthse des
travaux mardi 13 mars, lors dune
confrence de presse de lAgence
nationale de recherche sur le sida
et les hpatites virales (ANRS).
Ladministration dune combi-
naison dantirtroviraux rend
le VIH indtectable dans le sang.
Cependant, le virus na pas dis-
paru et persiste ltat latent
dans certaines cellules. Il est
notamment intgr au gnome
des lymphocytes T CD4 dans les
ganglions lymphatiques, lintestin
et le cerveau, qui constituent son
rservoir.
Courtney Fletcher (universit du
Nebraska) a fait part de donnes
accrditant lhypothse selon
laquelle une pntration ina-
dquate des mdicaments contri-
buerait une rplication occulte
du VIH, qui entretient linfection
par ce virus .
La taille de ce rservoir viral peut
tre contrle de telle manire
quelle demeure rduite. Cest ce
qui se produit chez les personnes
capables spontanment de conser-
ver linfection ltat latent. Il en
va de mme chez des patients mis
trs prcocement sous antirtro-
viraux, tels ceux de lessai ANRS
Visconti , ds la primo-infection
par le VIH (dans les quinze jours
ou les tout premiers mois suivant
la contamination).
Ractivation du virus
Les travaux prsents la CROI
ont montr quun traitement trs
prcoce diminuait fortement le
rservoir. Ce qui est cohrent avec
ce que nous avons observ chez les
patients de lessai Visconti : plu-
sieurs annes aprs linterruption
du traitement, qui avait dur en
moyenne trois ans, la charge virale
demeurait indtectable , indique
Christine Rouzioux.
Mais comment radiquer le VIH ?
Toute la difcult est que les
cellules dans lesquelles le VIH per-
siste ltat latent sont des lym-
phocytes CD4 dots de la mmoire
immunitaire et possdant une
longue dure de vie. Ces cellules
sont trs bien protges par le
systme immunitaire , souligne
Christine Rouzioux. Comme il est
inconcevable dliminer tous les
CD4, une autre stratgie est explo-
re. Sharon Lewin (Monash Uni-
versity, Melbourne) la prsente
Seattle : utiliser des agents pour
ractiver les cellules ltat latent.
Cela dclencherait nouveau la
rplication virale, et les antirtro-
viraux pourraient alors agir.
Plusieurs mdicaments employs
contre certains cancers, apparte-
nant la classe des inhibiteurs
dune enzyme, lhistone dacty-
lase (HDACi), ont t tests avec
succs sur un petit nombre de pa-
tients. Nous nensommes quaux
balbutiements. Surtout, il y a des
interrogations thiques majeures
recourir ces traitements lourds
chez des patients qui se portent
plutt bien , tempre Christine
Rouzioux.
Les chercheurs estiment indispen-
sable que pouvoirs publics et in-
dustriels simpliquent fortement
dans ce d majeur. La Socit in-
ternationale sur le sida (IAS), qui
rassemble la communaut scienti-
que dans ce domaine, et Franoise
Barr-Sinoussi, qui en prendra la
prsidence en juillet, lancent lini-
tiative HIV Cure , centre sur la
question des rservoirs viraux. Un
accord-cadre sur cette recherche va
tre sign entre plusieurs institu-
tions publiques, notamment
lANRS et les instituts nationauxde
la sant amricains.
Paul Benkimoun
(17 mars 2012)
POURQUOI
CET ARTICLE ?
Ces dernires annes, le d-
veloppement des trithrapies
contre le VIH est lorigine
dun progrs considrable
dans la prise en charge des
patients infects par ce virus.
La prise de traitements an-
tiviraux, dbute un stade
prcoce de linfection, retarde,
voire empche, le dveloppe-
ment du stade sida chez le
patient. Cependant, chez le
patient sropositif pour le
VIH et trait par trithrapie,
si la charge virale devient trs
faible voire infrieure au seuil
de dtection, le virus reste
prsent dans lorganisme. Le
patient est donc capable de
transmettre le virus. Larticle
prsente les pistes envisages
pourliminertotalement leVIH
delorganisme, enciblant les r-
servoirs de lorganisme abritant
certrovirus. Il sagit pour lins-
tant de pistes de recherches
approfondir. Si ces pistes
se rvlaient prometteuses,
elles pourraient contribuer
faire rgresser lpidmie,
alors quun vaccin efficace
tarde toujours tre mis sur
le march.
Une piste pour radiquer le VIH de lorganisme
De nouveaux traitements pourraient liminer les rservoirs viraux dans lesquels le
virus du sida persiste.
les cellules dendritiques , ces
fantassins delimmunitchargs de
prsenter les antignes auxcellules
immunes pour leur apprendre
tuer les porteurs de ces antignes.
Les cellules dendritiques sont
prleves chez les patients, mises
incuber avec des fragments de
molcules spciques de certains
cancers ( antignes tumoraux ),
puis rinjectes aux patients. Dans
les cancers de la prostate hormono-
rsistants mtastatiques, cette m-
thode approuve aux tats-Unis
permet des gains de survie de
six huit mois , observe Laurence
Zitvogel.
Autre voie : rinjecter au patient
ses propres lymphocytes aprs les
avoir activs exvivopar diffrentes
manipulations. Au pralable, le pa-
tient doit avoir subi une ablationde
ses cellules mdullaires. Ce sont
des protocoles trs lourds et co-
teuxmais assez rvolutionnaires ,
souligne la chercheuse. Ils ont fait
la preuve du concept dans des
mlanomes et des leucmies lym-
phodes chroniques.
Les tumeurs sont loin davoir sign
leur reddition. Mais face aux
charges ritres des escadrons de
limmunit, renforcs de ces nou-
velles armes thrapeutiques, elles
amorcent un recul manifeste.
Florence Rosier
(8 novembre 2012)
*Co-investigatrices des essais
cliniques avec lipilimumab, les
professeurs Caroline Robert et
Cleste Lebb dclarent des liens
dintrt avec les laboratoires
Bristol-Myers Squibb notamment.
POURQUOI
CET ARTICLE ?
Plusieurs stratgies thrapeu-
tiques existent pour lutter
contre les cancers : la chirurgie,
qui consisteliminer physique-
ment les tumeurs et les mtas-
tases ; la chimiothrapie, qui tue
les cellules cancreuses avec des
molculescytotoxiques; laradio-
thrapie, qui dtruit les cellules
cancreusesenlesexposant des
radiations, et, enn, limmuno-
thrapie. Limmunothrapievise
utiliser lesystmeimmunitaire
du patient pour quil limine
lui-mmeles cellules tumorales.
Le concept de limmunoth-
rapie est ancien mais na pas
encore totalement prouv son
efcacit thrapeutique. Actuel-
lement, certains traitements
interessants sont lessai. Cet
article dtaille les diffrentes
stratgies en cours d'tude en
immunothrapie : certaines
dentre elles semblent parti-
culirement prometteuses. La
progression des connaissances
de mcanismes cancreux
dune part et des mcanismes
immunitaires dautre part
devrait permettre damliorer
lapproche immunothrapique
du traitement des cancers.
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
78
LESSENTIEL DU COURS
Le maintien de lintgrit de lorganisme : quelques aspects de la raction immunitaire 79
LESSENTIEL DU COURS
Le maintien de lintgrit de lorganisme : quelques aspects de la raction immunitaire
Le phnotype
immunitaire au cours
de la vie
Le dbut de la vaccination
Les mcanismes de la raction inflammatoire et
de la mmoire du systme immunitaire ne sont
connus que depuis le XX
e
sicle, mais la raction
de lorganisme contre les pathognes avait dj
t observe et tudie de faon empirique.
Depuis lAntiquit, on a remarqu que les sur-
vivants dune pidmie pouvaient tre protgs
contre lpidmie suivante de la mme maladie.
Gurir dune maladie permet donc dtre protg
contre celle-ci des annes durant. Cependant,
cette constatation empirique est loin dtre sys-
tmatique : ainsi, on peut nouveau tre atteint
par le rhume, alors quon en avait guri lanne
prcdente. Pourquoi existe-t-il une mmoire
immunitaire contre certaines maladies mais pas
contre dautres ? La lutte contre la variole permit
de rpondre cette question. Avant le XVII
e
sicle,
les mdecins pratiquaient la variolisation, qui
consistait inoculer des patients sains de
vieilles pustules dessches de varioleux. Cette
pratique prsentait un risque mais permettait une
bonne protection contre une pidmie de cette
maladie mortelle. Un mdecin anglais, Edward
Jenner, a constat que les paysans au contact
des vaches contractaient une maladie bovine
proche de la variole : la vaccine, qui les protgeait
contre les pidmies de variole. En 1796, Jenner
eut lide (selon un protocole que lthique m-
dicale rprouverait aujourdhui) dinjecter un
jeune garon des extraits de pustules, prlevs
sur une jeune vachre atteinte par la vaccine. Il
inocula ensuite la variole au jeune garon, qui ne
contracta pas la maladie. Cette dcouverte lore
du XIX
e
sicle ouvrit la porte ce qui fut appele
la vaccination. Les travaux de Louis Pasteur sur
le cholra des poules et sur la rage ont confirm la
protection que linjection dun agent ressemblant
lagent pathogne confre lorganisme contre
ce mme agent pathogne.
Le principe de la vaccination
La premire rencontre dusystme immunitaire avec
un agent pathogne dclenche une rponse immu-
nitaire dite primaire qui consiste en la mise en
place de cellules effectrices (plasmocytes scrteurs
danticorps, lymphocytes Tauxiliaires, lymphocytes T
cytotoxiques) spciques de cet agent. Lors dun
contact ultrieur avec le mme agent infectieux, la
rponse immunitaire dite secondaire est plus
rapide et quantitativement plus importante. Eneffet,
la raction immunitaire primaire saccompagne de
la formation de cellules immunitaires mmoires
longue dure de vie : il sagit des lymphocytes T
et Bmmoires. Les tudes ont montr que certaines
cellules immunitaires, dont certains lymphocytes Bet
T, peuvent vivretrs longtemps, aumoins unedizaine
dannes, dans lorganisme. Ainsi, lors dun contact
ultrieur aveclagent infectieux, les LTet LBmmoires
ragissent beaucoup plus vite lagent infectieux
en prolifrant et en se diffrenciant beaucoup plus
rapidement que les LT et LB nafs, qui nont jamais
t activs par lagent infectieux. Lexistence de cette
ractionsecondaireintenseet rapidencessitequeles
LTet LBmmoires ragissent des antignes proches
de ceux les ayant activs la premire fois.
Leprincipeduvaccinutilisecettemmoireimmuni-
taire: lepatient est exposunpathognemodiet
nondangereux(pathogne tu, inactiv oumorceau
de pathogne) mais immunogne, cest--dire en-
tranant une raction immunitaire sufsante pour
former des LB et des LT mmoires.
Il est souvent ncessaire deffectuer des rappels
de vaccination pour entretenir cette mmoire
immunitaire. Ainsi, par la vaccination, le patient
qui na pas dvelopp la maladie cause par
lagent infectieux possde un phnotype im-
munitaire qui lui permet lors dun contact ult-
rieur avec lagent infectieux de dclencher une
rponse immunitaire secondaire, suffisament
rapide et intense, pour lui viter de tomber malade.
L
e phnotype immunitaire dun individu est sa capacit
rpondre aux agents infectieux quil rencontre. Or, cette
capacit de rponse nest pas ge dans le temps mais
volue au cours de la vie : certaines maladies ne peuvent pas
tre contractes deux fois et la vaccination confre une protec-
tion contre des infections. Comment expliquer lvolution du
phnotype immunitaire au cours de la vie ?
Ladjuvant, dclencheur
de la raction inne, indispensable la
raction adaptative
De nombreux vaccins contiennent, enplus de lagent
vaccinant, un adjuvant. Ladjuvant est une substance
(comme les sels daluminium ou le squalne) qui
dclenche une raction immunitaire inne, cest-
-dire une raction inammatoire. Cette raction
inammatoire, duelaprsencedeladjuvant, stimule
lactivationdes cellules dendritiques, qui phagocytent
lagent vaccinant et le dgradent en antigne. Ces
cellules dendritiques migrent vers les ganglions lym-
phatiques et prsentent lantigne expos au sein de
leur CMHauxlymphocytes TCD4+
. Ces lymphocytes T
auxiliaires activs par lantignestimulent ladiffren-
ciation des LB en plasmocytes scrteurs danticorps.
Ainsi ladjuvant duvaccindclenche la ractioninne
indispensable linstallation de la raction immu-
nitaire adaptative, dont lintensit est accrue. Le rle
des adjuvants prsents dans les vaccins souligne
limportancedelacooprationentrelimmunitinne
et limmunit acquise.
La vaccination et la sant publique
La vaccination a pour objectif une protection indivi-
duelle, mais aussi une protection collective, et elle
peut permettre une radication du pathogne,
condition que lhomme soit le seul hte de cet agent
pathogne. Cest le cas en particulier de la variole,
radique sur tous les continents depuis 1979. La
couverturevaccinaledunepopulationreprsentele
pourcentage de la population vaccine. On considre
quil faut environ une couverture vaccinale de 95 %
pour quunemaladiesoit absentedelapopulation. La
vaccination constitue donc un vritable enjeu de
sant publique.
Prol de productiondes anticorps lors dune immunisationet
structure des IgMet IgG
Attraction
des cellules
dendritiques
Cellule
dendritique
Activation des LB
Migration vers les
organes lymphodes
(rate, ganglions)
Mmoire
Plasmocyte
producteur
danticorps
Mmoire
Ativation Ativation
LB
LB
LTaux LTaux
LTaux
LTaux
LTCD4+
LTaux
LB
LB
LB
LB
LB
tapes dactivation du systme immunitaire par un vaccin
(LTaux =LTauxiliaire)
ZOOM SUR
LA DISPARITION
DES MALADIES
De nouvelles maladies appa-
raissent (sida, etc.), certaines
disparaissent, avec ou sans vacci-
nation. La peste, lorigine dpi-
dmies meurtrires au Moyen
ge en Europe, a disparu au
XVIII
e
sicle car les individus pr-
sentant une meilleure rponse
immunitaire contre la peste ont
t slectionns d'un point de
vue volutif. En octobre 1977,
lOrganisation mondiale de la
sant (OMS) a dclar totalement
radique une maladie virale
trs contagieuse : la variole ou
petite vrole . Cette radica-
tion est le rsultat de campagnes
de vaccination massive mises en
uvre partir des annes 1960
sur tous les continents.
Actuellement, seuls quelques la-
boratoires au monde sont autori-
ss conserver des chantillons
du virus de la variole. Cette
conservation fait dbat au sein
de la communaut scientique :
quel est lintrt de conserver
ce virus particulirement dan-
gereux pour les populations
actuelles qui ny ont jamais t
exposes ?
LE VACCIN DE PASTEUR
CONTRE LA RAGE
En 1885, un enfant, Joseph Meis-
ter, g de 9 ans, fut mordu par
un chien enrag : il consulta
rapidement Louis Pasteur, bien
avant que sa rage soit dclare.
Pendant une dizaine de jours,
Pasteur lui inocula une sus-
pension de moelle osseuse de
lapin mort de la rage et Joseph
Meister ne dveloppa pas cette
maladie mortelle. Les injections
ont permis son systme im-
munitaire de dvelopper une
rponse immunitaire primaire
avant que se dveloppe le virus.
Puis, la rponse immunitaire
secondaire a permis lenfant de
lutter contre le virus contract
lors de la morsure. Par la suite,
Pasteur t mme une injection
de lagent responsable de la rage
lenfant pour prouver la protec-
tion confre par la vaccination.
DATES CLS
Limmunologie
1796: Jennerpratiquelespremires
vaccinations contre la variole avec
la vaccine bovine.
1878 : mise au point par Pasteur
de la vaccination avec des agents
pathognes.
1882: dcouvertedelaphagocytose
par Metchnikoff.
1890: isolement des anticorps par
vonBehring et Kitasato.
1980 : identication des gnes co-
dants pour les LCR des LT et BCR
des LB.
1983 : identication du VIH par
lquipe de Montagnier.
2011 : prix Nobel de Mdecine
dcern Hoffmann, Beutler et
Steinmann pour leurs travaux sur
limmunit inne (rcepteurs Toll
et TLR, cellules dendritiques).
REPRE
Les vaccinations
Diphtrie, ttanos, coqueluche,
poliomylite (vaccination obliga-
toire) : 3 injections 2, 3 et 4 mois,
puis4rappelsavant 18ans. Rappels
tous les dixans chez ladulte.
Rougeole, oreillons, rubole : 1 in-
jection 12 mois, puis une 2
e
entre
13 et 24 mois.
Hpatite B : 3 injections 2, 4 et
16-18 mois.
Tuberculose : vaccination non
obligatoire pour lentre en col-
lectivit depuis 2007 mais recom-
mande pour les enfants risque
levdetuberculose(le-de-France,
Guyane, Mayotte, antcdents fa-
miliauxdetuberculose, etc.) : injec-
tion ds la naissance et injections
supplmentaires entre3et 11 mois,
et aprs 12 mois si ncessaire.
Grippe saisonnire : vaccination
recommande partir de 65 ans,
aux femmes enceintes, aux per-
sonnes avec diverses pathologies
(dont lobsit) : une injectiontous
les ans.
De nombreuses autres vaccina-
tions existent (coqueluche, vari-
celle, hpatite A, papillomavirus
humain, pneumocoque, mnin-
gocoque, etc.) La vaccination est
adapter chaque individu en
fonction de son lieu de vie, de son
modedevieet desontat desant.
DEUXARTICLES DU MONDE
CONSULTER
Vaccins Laluminium est-il nfaste ?
p. 81-82
(Stphane Foucart, 3 novembre 2012)
Vaccin contre lhpatite B p. 82
(P. Be, 18 fvrier 2012)
LesIgM, constitusdecinqimmunoglobulines, sont produitspar lesplasmocytesdont lerecrutement est rcent, tandisqueles
IgGsont produits par des plasmocytes dont lactivationest plus ancienne.
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
80
UN SUJET PAS PAS
Le maintien de lintgrit de lorganisme : quelques aspects de la raction immunitaire
LES ARTICLES DU
81 Le maintien de lintgrit de lorganisme : quelques aspects de la raction immunitaire
Lintitul complet du sujet
Entre 3 et 18 mois, un enfant a t admis de trs
nombreuses fois lhpital pour diverses infections
bactriennes graves. II areutous les vaccins (ttanos,
diphtrie, rougeole, etc.) prvus lgede2, 3et 4mois.
Des analyses ont t ralises et ont permis de dter-
miner quil tait atteint dune maladie hrditaire (la
maladiedeBruton). Exploitez les informations saisies
dans le document propos andexpliquer pourquoi
lorganismedecet enfant est incapabledelutter contre
les infections bactriennes.
Le document
Rsultats des examens de laboratoire ayant permis
le diagnostic
*Les anatoxines sont des protines bactriennes.
**Anticorpsetimmunoglobulinessontdessynonymes;
ces molcules sont produites par les lymphocytes B.
Proposition de corrig
Lenfant areutouslesvaccinsprvuslgeprconis,
mais les tests raliss montrent labsence danticorps
spciques dirigs soit contrelanatoxinediphtrique
(vaccinationcontreladiphtrie), soit contrelanatoxine
ttanique(vaccinationcontrelettanos), soit contrele
virus de la rougeole (vaccination contre la rougeole).
Lavaccinationnapasentranlapparitiondanticorps
dirigs spciquement contre les antignes contenus
dans les vaccins comme cela aurait d tre le cas. La
vaccinationnadonc pas teffectiveet ellenapas pu
confrer de protection.
Le taux dimmunoglobulines G de lenfant est trs
faible (0,17 g.L
-1
alors que le taux normal est de 5,5
10,0 g.L
-1
) et les immunoglobulines A ne sont pas
dtectes (leur taux normal est de 0,3 0,8 g.L
-1
). Cet
enfant produit donc trs peu dimmunoglobulines,
ce qui explique ses graves infections microbiennes
rptes et linefcacitdelavaccination.
Enfin, ltude des populations lym-
phocytaires de lenfant montre que
le nombre de lymphocytes totaux est
normal chez cet enfant (3,05.10
7
L
-1
pour
un taux normal de 2,5.10
7
L
-1
5.10
7
L
-1
).
Mais si le taux de LT est dans la norme
(3,02.10
7
.L
-1
pour un taux normal de
1,5.10
7
L
-1
3,0.10
7
L
-1
), le taux de LB de
0,03.10
7
.L
-1
est trs faible, reprsentant
environ un tiers de la valeur minimale.
Ainsi cet enfant prsente une anomalie
importante du nombre de LB. Son trs
faible taux de LB ne lui permet pas de
produire assez danticorps pour lutter
efcacement contre des infections bac-
triennes. De plus, sa carence enLBrend
inefcace toute vaccination: il est donc
susceptible de contracter des maladies
que les autres enfants vaccins ne d-
clarent pas. Cette trs faible quantit de
LB dans son organisme a une origine
hrditaire.
Partie 2.1 :
Diagnostic immunitaire
chez un enfant
Vaccins reus
Dosage des anticorps** spcifiques
en rponse aux vaccinations
Dosage des immunoglobulines**
du srum chez le patient
Valeurs normales
pour lge de 18 mois
Sous-population lymphocytaire
du sang
Pas danticorps spcifiques
dtects
Pas danticorps spcifiques
dtects
Pas danticorps spcifiques
dtects
0,17 g.L-1 5,5 10,0 g.L-1
3,05.107 L-1 2,5.107 g.L-1 5.107 g.L-1
0,03.107 L-1 0,1.107 g.L-1 0,4.107 g.L-1
3,02.107 L-1 1,5.107 g.L-1 3,0.107 g.L-1
Non dtectes 0,3 0,8 g.L-1
Anatoxine* ttanique
Anatoxine diphtrique
Virus de la rougeole
Immunoglobulines G
Immunoglobulines A
Lymphocytes totaux
Lymphocytes B
Lymphocytes T
Valeurs normales
pour lge de 18 mois
Daprs H. Chapel, M. Haeney, S. Misbah, N. Snowden, Immunologie
clinique
Ce quil ne faut pas faire
Se contenter de saisir les donnes du document
sans les mettre en relation et sans expliquer
lorigine des infections bactriennes rptes.
AUTRES SUJETS POSSIBLES AU BAC
SUR CE THME
tude de documents
Mise envidence de la mmoire immunitaire.
Justicationducalendrier de vaccination.
tude de mise au point de vaccins (VIH, trypano-
some, paludisme, vaccinanti-tumeur, etc.).
tude durle des adjuvants.
ZOOM SUR
VACCIN ANTI-VIH :
DES PISTES, MAIS
UN VACCIN EFFICACE
TOUJOURS ATTENDU
Lisolement du VIH en 1983 avait
ouvert la perspective de lobten-
tion rapide dun vaccin. Mais
lextrme variabilit du VIH a
jusqu maintenant rendu dif-
cile la mise au point dun vaccin.
Les traitements antirtroviraux
(comme le Truvada) ont rcem-
ment montr quils rduisaient le
risque de transmissionduVIHpar
voie sexuelle, mais la condition
dtre pris quotidiennement. En
2009, des essais cliniques d'un
vaccin anti-VIH ont montr une
efficacit partielle (31 %) mais
intressante. Les chercheurs
progressent actuellement dans
lexplication de ces rsultats et
esprent pouvoir amliorer lef-
cacit de ce vaccin. Actuellement,
aux tats-Unis, lefcacit dun
autre vaccin ayant montr son
innocuit est en cours dtude
chez lhomme.
CANCERS ET
IMMUNOTHRAPIE
L immunothrapie passive
consiste administrer au ma-
lade des anticorps articiels dits
monoclonaux , ayant une cible
molculaire prcise la surface
des cellules cancreuses dans
lobjectif de dtruire ces der-
nires. Des anticorps contre des
facteurs qui interviennent dans
la multiplicationdes cellules sont
galement utiliss pour bloquer la
prolifration cellulaire.
Limmunothrapie active vise
mobiliser ou renforcer les res-
sources du systme immunitaire
pour accrotre llimination des
cellules cancreuses. Elle consiste
stimuler lactivit globale du
systme immunitaire du malade
en utilisant des cytokines (inter-
fron par exemple). Des cellules
tumorales du patient peuvent
tre cultives pour les rendre
plus immunognes avant de
les rinjectes. Enn les cellules
immunitaires peuvent tre modi-
es pour les rendre plus efcaces
contre les cellules tumorales.
Vaccins Laluminium est-il nfaste ?
La vaccination sauve des millions de vies. Mais un adjuvant rpandu est souponn
dengendrer divers syndromes chez des personnes ayant un prol gntique particulier.
D
ouleurs musculaires par-
fois intenses, troubles co-
gnitifs, fatigue profonde
et invalidante que le sommeil ne
parvient pas rparer quoi
sajoute le dsarroi de ne pas
comprendre. Voici les principaux
symptmes associs une nou-
velle entit pathologique quun
groupe de mdecins conduits par
Romain Gherardi (hpital Henri-
Mondor, Inserm, universit Paris-
Est) et Michelle Coquet (hpital
Pellegrin, Bordeaux) dcrivaient,
en aot 1998, dans The Lancet.
lpoque, cette affectiondorigine
inconnue, baptise myofasciite
macrophages , reposait sur des
observations menes sur une pe-
tite vingtaine dindividus. Depuis,
la cohorte du centre de rfrence
des maladies neuromusculaires
de lhpital Henri-Mondor (Cr-
teil) a grandi, avec prs de 600pa-
tients suivis aujourdhui.
Au sein de la petite communaut
de chercheurs et de praticiens
qui travaillent sur le sujet, un
consensus merge : la myofasciite
macrophages serait provoque
par lhydroxyde daluminium
utilis dans la plupart des vac-
cins. Mais cette attribution est
prement discute. Aucun lien
de causalit na t clairement
dmontr entre la vaccination et
la myofasciite macrophages ,
dit ainsi limmunologiste Jean-
Franois Bach, ancienmembre du
comit sur la scurit des vaccins
de lOrganisation mondiale de la
sant (OMS). Lpidmiologie na
pas encore permis de trancher.
Une chose est cependant sre.
Les biopsies pratiques dans le
muscle deltode (cest--dire sur
le site des injections vaccinales)
des individus atteints de ce
syndrome montrent des lsions
spciques, avec des nanocris-
taux daluminium pigs dans
les macrophages ces cellules
qui participent limmunit en
phagocytant les corps trangers
ou les dbris de cellules mortes.
Chez lcrasante majorit des
individus, ces lsions persistent
quelques jours quelques se-
maines aprs la vaccination, mais
chez les personnes atteintes de
myofasciite, elles persistent de
trs nombreuses annes.
Les adjuvants vaccinaux lalumi-
niumsont utiliss depuis presque
un sicle. Ils permettent de crer
une rponse immunitaire sans
laquelle le vaccinserait inefcace
mais sont, ces dernires annes,
au centre de polmiques rcur-
rentes. En mars, un groupe de
dputs a mme propos un mo-
ratoire sur laluminiumvaccinal
Pour RomainGherardi, le lienfait
peu de doute entre ces adjuvants
et le syndrome quil a contribu
identier en 1998. Ce que de-
vient laluminium vaccinal dans
lorganisme na presque pas t
tudi : tout repose sur le postulat
quil se dissout dans le liquide
interstitiel avant dtre vacu,
dit-il. En ralit, les seules exp-
riences ont t faites sur deux
lapins pendant vingt-huit jours
[publies en 1997 dans la revue
Vaccine]. Ces rsultats ont t ex-
trapols lhomme, sans aucune
tude sur le long terme.
Les observations menes sur
les personnes atteintes de myo-
fasciite suggrent que les sels
daluminiumne se dissolvent pas
spontanment mais quils sont
phagocyts par les macrophages.
Chez la plus grande part de la
population, ces cellules-boueurs
engloberaient ces nanocristaux
daluminiumet les soumettraient
une acidit sufsante pour les
solubiliser, permettant ainsi lor-
ganisme de sen dbarrasser plus
oumoins rapidement. Mais, sur
certains terrains gntiques, les
macrophages qui pigent lalumi-
nium ne parviennent pas nir
le travail, assure M. Gherardi. Or,
tant quun macrophage nest pas
parvenu se dbarrasser de la
particule quil a phagocyte, il
devient immortel. Il rejoint la cir-
culationsanguine et a une proba-
bilit accrue de passer la barrire
hmato-encphalique pour se
retrouver stock dans le cerveau,
dont il ne ressortira pas.
Cette persistance des particules
daluminium dans les cellules
immunitaires dissmines dans
lorganisme provoquerait une
raction inflammatoire chro-
nique responsable de douleurs
musculaires et de troubles cogni-
tifs, signant lexposition accrue
des tissus nerveux laluminium.
Ce tableau serait donc celui de
la myofasciite macrophages.
Combien de personnes seraient
effectivement touches ? Impos-
sible de le savoir prcisment ,
dit M. Gherardi.
Pour M. Bach, rien de cela nest
dmontr. Il est indniable
quune petite part de lalumi-
nium qui entre dans lorganisme
pntre dans le cerveau, prcise
limmunologiste, mais ce sont des
quantits minimes Une dose de
vaccin contient moins dalumi-
nium que ce que nous ingrons
par voie alimentaire. Ce quoi
M. Gherardi rpond que lalumi-
nium particulaire ingr na que
trs peu de chances de nir sous
cette forme dans la circulation
sanguine, au contraire de celui
qui est inject.
Le dbat est vif : lexpression
dun dbat scientique sur des
risques possibles de la vaccina-
tion pour une petite fraction des
individus pourrait en dtourner
le plus grand nombre, exposant
du mme coup lensemble de la
population des dangers bien
plus grands. La questionouvre en
outre de nouveaux champs de re-
cherche. En2011, limmunologiste
Yehuda Shoenfeld, professeur
luniversit de Tel-Aviv (Isral), a
propos dans Journal of Autoim-
munity de rassembler une srie
de pathologies sans lien appa-
rent sous le terme de syndrome
auto-immunitaire induit par des
POURQUOI
CET ARTICLE ?
La faible participation la cam-
pagne de vaccination contre le
virus H1N1 en 2008 a montr
les rticences dune partie de
la population franaise se
faire vacciner. Un des argu-
ments souvent utilis contre
la vaccination fut la dange-
rosit associe ladjuvant
utilis : les sels daluminium.
Ces sels sont accuss dtre
responsables dune patholo-
gie appele myofasciite
macrophages . Lors de la vac-
cination, les sels daluminium,
en dclenchant la rponse
immunitaire inne, potentia-
lise leffet du vaccin mais les
particules daluminium, pha-
gocytes par les macrophages,
resteraient longtemps dans
ces cellules immunitaires et
seraient lorigine de troubles
affectant diffrents tissus, dont
le tissu nerveux. Larticle fait le
point sur ltat des recherches
sur le devenir de laluminium
dans lorganisme suite une
vaccinationet met galement en
lumire la difcult dtablir un
lien de causalit ou dabsence
de causalit entre laluminium
et les troubles associs par les
tudes pidmiologiques.
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
82
LES ARTICLES DU
NEURONE ET FIBRE
MUSCULAIRE :
LA COMMUNICATION NERVEUSE
Deglutition
Langue
Mchoire
Lvres
V
isage
il
S
o
u
r
c
i
l
C
o
u
P
o
u
c
e
D
o
i
g
t
s
M
a
i
n
P
o
i
g
n
e
t
C
o
u
d
e
B
r
a
s
p
a
u
l
e
Aire
motrice
primaire
Orteils
Cheville
G
e
n
o
u
H
a
n
c
h
e
T
r
o
n
c
C
ette controverse fait de la
France une exception : des
rticences la vaccination
contre lhpatite Bpersistent, bien
que les tudes naient pas tabli
lexistence dune association sta-
tistiquement signicative avec la
survenue dune affection de type
sclrose en plaques (SEP).
Lance en 1994, la campagne de
vaccination devait concerner les
nourrissons et les enfants gs de
10 11 ans. Enraisondundiscours
alarmiste, notamment des fabri-
cants de vaccins, elle a massive-
ment t tendue aux adultes et
la tranche dge la plus susceptible
de dvelopper une SEP.
Aucun dispositif spcifique de
suivi prospectif nayant t prvu
par les autorits sanitaires, la
seule possibilit pour tenter den
avoir le cur net tait de raliser
une tude rtrospective des cas-
tmoins.
Lorsque nous avons prsent
nos rsultats Bernard Kouch-
ner, secrtaire dtat la Sant en
1998, il nous a dit : En quelque
sorte, vous me mettez dans la
merde. Nous avions en effet re-
trouv un excs de risque de SEP
chez les personnes vaccines mais
qui ntait pas statistiquement
signicatif et ne permettait pas
de conclure , se souvient Ber-
nard Bgaud (universit Bordeaux
Segalen).
Les diffrentes tudes effectues
ne mettent pas envidence de lien.
Elles ne signient pas pour autant
quil soit excluque le vaccinprci-
pite lapparition dune SEP sous-
jacente. Cette affaire illustre bien
que, mme si lon employait la
meilleure mthodologie du
monde, lpidmiologie ne peut
pas rattraper le fait que lon ne se
soit pas organiss pour disposer de
donnes de suivi beaucoup plus
exhaustives , souligne Bernard
Bgaud.
P. Be
(18 fvrier 2012)
Vaccin contre lhpatite B
POURQUOI
CET ARTICLE ?
Lhpatite B est une infection
virale qui, dans environ un cas
sur dix, est lorigineduncancer
du foie. Le virus de lhpatite B
se transmet par voie sexuelle ou
sanguine. Levaccinexistedepuis
1982 et confre une protection
efcace. Il est mme obligatoire
pourcertainsmtiers, commeles
professionnels de la sant. Mais
en France, des cas de sclrose en
plaques, une maladie auto-im-
mune dans laquelle le systme
immunitaire dtruit les propres
cellules de lorganisme, semblent
tre apparus aprs vaccination
contre lhpatite B. Cette suspi-
ciondecorrlationnest observe
quen France. Cet article fait le
point de manire synthtique
sur lavaccinationcontrelhpa-
titeB. Lacausalitentrelasclrose
enplaquesetlavaccinationcontre
lhpatiteBnapastdmontre,
car les tudes pidmiologiques
ne permettent pas de conclure
avec certitude. Le sujet demeure
controvers, de par labsence de
rsultats scientiques clairs, et
sesttraduitparunebaisseimpor-
tante de la couverture vaccinale
contre lhpatite B ces dernires
annes enFrance.
adjuvants ( ASIA en anglais).
Avec, au ct de la myofasciite, le
syndromedelaguerreduGolfe, at-
tribuparplusieurschercheursaux
nombreux vaccins adjuvs lalu-
minium ou au squalne. Ce syn-
drome est lui aussi caractris par
une fatigue intense, des troubles
cognitifs, desdouleursmusculaires
et articulaires chroniques
Une grande part de ce qui est
rassembl dans cet article est
compltement spculatif , dit
M. Bach. Consult par Le Monde,
un immunologiste franais,
tranger la controverse, juge
aussi svrement larticle, esti-
mant que lauteur fait de la
communication en tirant des
analogies sans rien expliquer .
En moins de dix-huit mois, le pa-
pier de M. Shoenfeld a cependant
t cit dans plus de cinquante
travaux ultrieurs.
Une autre pathologie le syn-
drome de fatigue chronique (SFC)
ou encphalopathie myalgique
touche auxtats-Unis environun
million de personnes et est, elle
aussi, caractrise par le mme
type de troubles. En 2009, la com-
munautscientiqueavait suivi la
piste infectieuse pour lexpliquer.
Mais laffaire sest solde n 2011
par un asco le rtrovirus sus-
pect, leXMRV(Xenotropic murine
leukemiavirus-relatedvirus) stant
rvltreunechimreaccidentel-
lement cre enlaboratoire.
Du coup, le SFC pourrait-il aussi
appartenir la famille ASIA ? Il
est trs concevable que de nom-
breux cas de fatigue chronique
qui sont aujourdhui dorigine
inconnue puissent tre lis, en
partie au moins, aux adjuvants ,
estime M. Shoenfeld. Mais aussi,
plusgnralement, lintroduction
de certains lments particulaires
danslorganisme(selsdaluminium
prsents dans certains cosm-
tiques, etc.). Selon M. Shoenfeld,
les ractions varient suivant le
terrain gntique. On a ainsi vu
apparatre des cas de narcolepsie
en Finlande aprs la campagne de
vaccinationcontreleH1N1 et aucun
en Allemagne, o cest pourtant le
mme vaccin [sans aluminium,
mais avec du squalne] qui a t
utilis. De fait, en mars, dans la
revue PLoS One, Markku Partinen
(universit dHelsinki) et ses coau-
teurs ont not une multiplication
par 17 de lincidence de la narco-
lepsie chez les enfants et les ado-
lescents en 2010, par rapport la
moyenne des annes prcdentes.
Sur 54nouveauxcas diagnostiqus
cetteanne, 50avaient tvaccins
contreleH1N1. Les premiers symp-
tmes tant apparus jusqu huit
mois aprs la vaccination.
Ces dlais rendent difcile, lors
dune simple consultation, lat-
tribution du syndrome la vac-
cination. De manire analogue,
selon M. Gherardi, les premiers
signes de la myofasciite peuvent
apparatre jusqu unanaprs la
dernireinjection. Autreobstacle
une meilleure comprhension
de ces maladies nouvellement
identifies : les techniques de
diagnostic, qui varient suivant les
pays. EnFrance, nous pratiquons
les biopsies musculaires dans le
muscle deltode, cest--dire l o
sont injects les vaccins, explique
M. Gherardi. Alors que, dans les
pays anglo-saxons, ces biopsies
sont pratiques loin du site din-
jection, dans le biceps, o lon ne
voit pas les lsions
Enn, et pour viter toute rcup-
rationde sontravail, M. Shoenfeld
veut conclure en rappelant que
la vaccination est ce que la m-
decine a donn de meilleur
lhumanit aucours des trois der-
niers sicles . Les entreprises
pharmaceutiques sont elles aussi
prudentes. Contactpar Le Monde,
Sano-Pasteur dit ne pas avoir de
porte-parole disponible pour
sexprimer sur la question et ren-
voie vers Les Entreprises du m-
dicament (LEEM), le syndicat du
secteur, qui lui-mme renvoie
vers le dernier rapport sur le sujet
de lAcadmie de mdecine selon
lequel lanalyse des conditions
ncessaires la provocationdune
maladie auto-immune napporte
aucune preuve ce jour permet-
tant dincriminer les vaccins ou
les adjuvants .
Stphane Foucart
(3 novembre 2012)
Le maintien de lintgrit de lorganisme : quelques aspects de la raction immunitaire
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
84
LESSENTIEL DU COURS
Neurone et bre musculaire : la communication nerveuse 85
LESSENTIEL DU COURS
Neurone et bre musculaire : la communication nerveuse
Le rexe myotatique,
un exemple de commande
rexe du muscle
Le rexe myotatique :
un outil diagnostique
Afin de vrifier ltat de tension qui sexerce sur
les muscles pour sopposer laction de la gravit
sur le corps humain, cest--dire pour tester le
tonus musculaire, le mdecin peut frapper avec
un marteau appropri soit au niveau du tendon
dAchille (au-dessus du talon), soit au niveau de
la rotule (sous le genou) et observer le rsultat.
Le rflexe myotatique est une raction rapide et
involontaire du corps. En raction son propre
tirement, le muscle se contracte automatique-
ment.
Diverses expriences historiques ont permis de
comprendre le mcanisme du rflexe myotatique.
Chez un chat dont la moelle pinire est section-
ne en-dessous de lencphale, le pincement de
la plante du pied dclenche un rflexe de retrait,
impliquant une contraction du muscle. Mais le
chat ne peut pas commander volontairement
le mouvement de sa patte. Ainsi le rflexe myo-
tatique ne fait pas intervenir le cerveau. Des
expriences de sections des nerfs reliant les
muscles la moelle pinire et de stimulations
aprs sections montrent que la moelle pinire
est le centre nerveux des rflexes myotatiques.
Ainsi, en tapant avec le marteau sur le tendon,
le mdecin tire artificiellement ce dernier : ce
stimulus va tre peru par des rcepteurs lti-
rement. Le message nerveux remonte via un
neurone sensoriel du nerf rachidien qui rejoint,
au niveau de sa racine dorsale (ou postrieure), la
moelle pinire. Les corps cellulaires des neurones
sensoriels sont situs dans les ganglions rachidiens
R
ester debout, ne pas perdre lquilibre, maintenir notre
posture malgr la pesanteur ncessitent un fonctionne-
ment permanent et adapt de nos muscles extenseurs
et chisseurs. Les muscles impliqus dans la posture comme
dans le mouvement sont des muscles stris squelettiques, atta-
chs au squelette par des tendons. Le mouvement des muscles
se rpercute sur le mouvement du squelette au niveau des ar-
ticulations. Dans le cas de la posture, ces mouvements ne sont
pas commands consciemment, ce sont des rexes muscu-
laires. Comment les rexes myotatiques commandent-ils la
contraction de certains muscles ?
1. Stimulus
2. tirement dumuscle
3. Perceptionde
ltirement
Synapse
neuromusculaire
Rcepteur sensoriel =
fuseauneuromusculaire
4. Transmissionde
linformation
Ganglionspinal
Racinedorsaledelamoellepinire
Substancegrise
Substanceblanche
5. Contractiondu
muscle
F
lu
x
d
in
f
o
r
m
a
tio
n
Tendon
Racineventraledelamoellepinire
Fibre nerveuse sensitive (afrente)
Fibres nerveuses motrices =
motoneurones (efrentes)
Circuit nerveuxdurflexemyotatique
et leurs extrmits sont en contact, au niveau de la
substance grise de la moelle pinire, avec le corps
cellulaire dun motoneurone ou neurone moteur.
Cette zone de contact entre les deux neurones est
appele synapse. Le message nerveux est alors
transmis du neurone sensoriel au motoneurone.
Les motoneurones cheminent par les racines
ventrales (ou antrieures) des nerfs rachidiens
jusquaux organes effecteurs que sont les muscles.
La zone de contact entre le motoneurone et le
muscle est appele synapse neuromusculaire ou
plaque motrice. La stimulation du motoneurone
entrane la contraction du muscle.
Le trajet du message nerveux du rcepteur sen-
soriel jusqu lorgane effecteur constitue un arc
rflexe, qualifi de monosynaptique. En effet,
durant tout le trajet nerveux, il nexiste quune
seule synapse, situe dans la substance grise de
la moelle pinire, entre le neurone sensoriel et
le motoneurone.
Le message nerveux
Toute cellule est caractrise par un potentiel
membranaire de repos, dont la valeur est de
70 mV. Dans certaines cellules dites excitables
comme les neurones, cette valeur du potentiel
membranaire peut tre modifie. Si la stimulation
du neurone est suffisante, elle entrane la modi-
fication transitoire de ce potentiel membranaire,
qui constitue un potentiel daction membranaire.
Un potentiel daction correspond une inversion
transitoire du potentiel du neurone. Le potentiel
daction est toujours identique : il a toujours la
mme dure et la mme amplitude. Il se propage
le long de laxone du neurone de proche en proche
sans sattnuer. Un message nerveux consiste en
une succession de potentiels daction transmis le
long de laxone dun neurone. Le message nerveux
est donc un message de nature lectrique dont
lintensit est code en frquence de potentiels
daction. Plus le nombre de potentiels daction
vhiculs par le neurone par unit de temps est
lev, plus lintensit du message nerveux est
forte.
Du motoneurone la cellule
musculaire : la synapse
neuromusculaire
Au niveau de la synapse neuromusculaire, zone de
contact entrelemotoneuroneet lemuscle, lemessage
nerveux de nature lectrique propag dans le moto-
neurone est traduit auniveaude la fente synaptique
en un message nerveux de nature chimique. Des
neuromdiateurs, ici l'actylcholine, scrts par le
motoneuronesexent sur des rcepteurs spciques
prsents la surface de la cellule musculaire. Cette
xationdclenche unpotentiels dactionmusculaire
lorigine de la contraction du muscle.
Ainsi, alors quau niveau dun neurone, lintensit
dumessage nerveux, de nature lectrique, est code
en frquence de potentiel dactions, au niveau de
la synapse neuromusculaire, le message nerveux
est de nature chimique et son intensit est code
en concentration de neuromdiateurs prsents
dans la fente synaptique.
Circuit nerveux dureexe myotatique Le potentiel daction: unmessage nerveux lmentaire
1
2 3
4
5
vsicule
actylcholine
cellule musculaire
acthylcholine
rcepteur
acthylcholine
UN ARTICLE DU MONDE CONSULTER
La moelle pinire, gardienne de la
locomotion automatique p. 87
(Herv Morin, 26 septembre 2009)
NOTIONS CLS
NEUROMDIATEUR,
NEUROTRANSMETTEUR
Molculelibreparunneuroneau
niveaud'unesynapseet capablede
transmettre le message nerveux
la cellule post-synaptique.
SYNAPSE
NEUROMUSCULAIRE
galement appele jonction
neuromusculaire ou plaque
motrice , la synapse neuromus-
culaire est la zone de contact entre
la terminaison synaptique dun
motoneurone et une cellule mus-
culaire. Larrive dun message
nerveux dans le motoneurone d-
clenche la contraction de la cellule
musculaire.
ZOOM SUR
LE SYSTME NERVEUX
Ensembledecellules, les neurones,
qui transmettent linformation
nerveuse dans lorganisme. Chez
les vertbrs, ondistingue :
le systme nerveux central, qui
comprend le cerveau et la moelle
pinire. Il est form de diffrents
centresnerveuxrelisentreeuxpar
des faisceaux de bres et contrle
lensemble dusystme nerveux;
lesystmenerveuxpriphrique,
qui est form de nerfs reliant la
priphrie de lorganisme et le sys-
tme nerveuxcentral.
LA MOELLE PINIRE
Centre nerveux situ sous
lencphale appartenant au
systme nerveux central. Dans
la moelle pinire, la substance
blanche, priphrique, correspond
aux axones des neurones, tandis
que la substance grise, centrale,
contient les corps cellulaires des
neurones. Elle est recouverte par
des membranes, les mninges ,
et abrite en son centre le canal de
lpendyme. Auniveaudes racines
dorsales (ou postrieures) de la
moelle pinire arrivent les bres
nerveuses sensorielles, dont les
corps cellulaires des neurones
sont contenus dans les ganglions
rachidiens. Au niveau des racines
ventrales (ou antrieures) sortent
les bres nerveuses motrices.
NOTIONS CLS
MESSAGE NERVEUX
Succession de potentiels dac-
tion qui se propagent le long des
neurones. Lintensit du message
nerveux est code en frquence
de potentiels daction.
MOTONEURONE
Neurone conduisant le message
nerveuxde la moelle pinire vers
un muscle.
NEURONE
galement appel cellule ner-
veuse , il est constitue dun
corps cellulaire et de prolonge-
ments, laxone et les dendrites.
Le corps cellulaire, galement
appel soma , est la partie
du neurone contenant le noyau.
Les dendrites sont les prolonge-
ments du neurone qui reoivent
des messages nerveux tandis que
laxone conduit le message ner-
veuxvers les terminaisons synap-
tiques. Unneurone est une cellule
excitable (capable de rpondre
un stimulus) et constitue lunit
de structure et de fonction du
systme nerveux.
POTENTIEL DACTION
Squence strotype de la mo-
dication du potentiel de mem-
brane dune cellule excitable. Le
potentiel daction est constitu
dune dpolarisation(le potentiel
daction atteint + 30 mV), suivie
dune repolarisation puis dune
hyperpolarisation transitoire
avant de revenir la valeur du
potentiel de repos ( 70 mV). La
dure du potentiel daction est
de quelques millisecondes. Le po-
tentiel dactionconstitue le signal
lmentaire dumessage nerveux.
POTENTIEL
DE MEMBRANE
Diffrence de potentiel lectrique
entre lintrieur et lextrieur de
la membrane plasmique de toute
cellule. Le potentiel membranaire
dit de repos est le potentiel de
membrane dune cellule excitable
en labsence de stimulation et sa
valeur est de 70 mV (lintrieur
de la cellule est plus ngatif que
lextrieur de la cellule).
Fonctionnement de la synapse neuromusculaire
Le message nerveux se propage sous forme de potentiels
daction le long du motoneurone (1) jusqu la terminaison
synaptique. Des vsicules charges dactylcholine
librent, par exocytose, leur contenu en neuromdiateurs
dans la fente synaptique (2). La xation de lactylcholine
sur ses rcepteurs localiss sur la membrane de la cellule
musculaire (3) gnre unpotentiel dactionmusculaire(4).
Ce potentiel daction musculaire dclenche la contraction
du muscle (5). Lactylcholine est ensuite dgrade dans
la fente synaptique et des molcules sont rinternalises
par la terminaison synaptique du motoneurone tandis que
la membrane est recycle par endocytose
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
86
UN SUJET PAS PAS
Neurone et bre musculaire : la communication nerveuse 87 Neurone et bre musculaire : la communication nerveuse
Lintitul complet du sujet
La myasthnie est une maladie caractrise par une
faiblesse musculaire : les patients narrivent pas
contracter efcacement leurs muscles. Exploitez len-
semble des rsultats exprimentaux pour proposer
une hypothse expliquant la myasthnie.
Le document
Exprience 1
Sur un muscle dun sujet sain et sur celui dun sujet
myasthnique, des lectrodes rceptrices sont places
proximit des synapses neuromusculaires. On
enregistre alors les potentiels dactions des cellules
musculairessuiteunestimulationdumotoneurone.
Exprience 2
L-bungarotoxineest unemolculetoxiquedevenin
de serpent, capable de se xer sur des rcepteurs
actylcholine. Linjection d-bungarotoxine une
souris la rend incapable de contracter ses muscles.
Exprience 3
On tudie par autoradiographie la fixation de
l-bungarotoxine radioactive sur les cellules muscu-
laires dun sujet sain et dun sujet myasthnique ; le
rsultat est prsent dans le tableau suivant :
Proposition de corrig
Dans lexprience 1, la stimulation du motoneurone
dclenche un message nerveux transmis le long du
motoneurone jusqu la synapse neuromusculaire,
zone de jonction entre le motoneurone et le muscle.
Chez le sujet sain, la stimulation du motoneurone
entraneuntrainsoutenudepotentiels dactionmus-
culaires (environ6/ms). Chez le sujet myasthnique,
cette stimulation provoque un train de potentiels
dactionmusculaires de la frquence plus faible (2/s).
Chez le sujet malade, le message nerveuxne parvient
donc pas correctement jusquau muscle.
L-bungarotoxine injecte la souris lempche de
contracter ses muscles. Cette toxine dclenche des
symptmes similaires ceux de la myasthnie. Or,
l-bungarotoxine se xe sur les rcepteurs ac-
tylcholine de la synapse neuromusculaire. On met
donc lhypothse que l-bungarotoxine en se xant
sur les rcepteurs actylcholine empche celle-ci
de sy xer : le message nerveux ne franchit pas la
synapse, do labsence de contraction des muscles
de la souris. La myasthnie serait donc due un
dysfonctionnement de la transmission du message
nerveux au niveau de la synapse neuromusculaire.
Daprs lexprience3, ladensitdel-bungarotoxine
xe sur la cellule musculaire est moins importante
chez le sujet myasthnique que chez le sujet sain. On
en dduit que la quantit de rcepteurs capables de
xer des molcules comme l'-bungarotoxine et
lactylcholine est plus faible chez le sujet myasth-
niquequechezlesujet sain. Ainsi lamyasthnieserait
due unnombre insufsants de rcepteurs pouvant
xer leur neuromdiateur : le message nerveux du
motoneurone nentrane pas alors une contraction
efcace du muscle.
Partie 2.1 :
La myasthnie
SUJET SAIN SUJET MYASTHNIQUE
30
0
-90
1 2 3
Temps (en s)
Potentiels daction
musculaires (en mV)
30
0
-90
1 2 3
Temps (en s)
Potentiels daction
musculaires (en mV)
sujet myasthnique sujet sain
+++ +
densit de
l -bungarotoxine
radioactive fixe
sur les cellules
musculaires
Ce quil ne faut
pas faire
Proposer une hypothse
sans justication
sappuyant sur les rsultats
exprimentaux.
AUTRES SUJETS POSSIBLES AU BAC SUR CE THME
Matrise des connaissances avec questionde synthse
Montrer comment letest dunrexemusculaireest unoutil dediagnosticdes anomalies dufonctionnement du
systme neuromusculaire. La synthse sera accompagne dunschma prsentant le rexe myotatique.
ZOOM SUR
Les molcules agissant
sur les synapses
De nombreuses substances
naturelles ou de synthse
peuvent agir sur les synapses
neuromusculaires et avoir
une action biologique sur les
muscles. Le mode daction de
ces molcules sur ces synapses
est variable, comme lillustrent
les exemples suivants.
Le curare est un poison v-
gtal obtenu partir de lianes
dAmrique du Sud, o il est
utilis, comme poison pour
chasser par certaines tribus. Les
pointes de flches enduites de
curare entranent la paralysie
de lanimal atteint. Comment le
curare dclenche-t-il de graves
paralysies musculaires ? Le
curare se fixe la place de lac-
tylcholine sur les rcepteurs de
la synapse neuromusculaire et
empche le message nerveux
de parvenir aux muscles. Le
curare est donc un antagoniste
de lactylcholine. Il est utilis
en mdecine pour ses effets
anesthsiants permettant le
relchement des muscles.
La nicotine se fixe sur les
rcepteurs actylcholine et
mime laction de lactylcho-
line. Elle est qualifie d ago-
niste de lactylcholine.
Le sarin est un gaz mortel,
qualifi de neurotoxique ,
qui dclenche une puissante
contraction des muscles, cest-
-dire une paralysie totale. Au
niveau de la synapse neuro-
musculaire, l actylcholine
fixe sur les rcepteurs de la
cellule musculaire est norma-
lement limine par une en-
zyme appele actylcholine
estrase . Or, le sarin inhibe
cette enzyme : lactylcholine
reste constamment fixe sur
son rcepteur, provoquant la
contraction permanente des
muscles. Le gaz sarin fut d-
couvert en Allemagne en 1939 ;
sa production et sa possession
sont interdits par lONU depuis
1991. Depuis 2007, plus aucun
pays ne devrait dtenir ce gaz
neurotoxique.
S
aint Denis, premier vque
de Paris, fut capable, dit sa
lgende, de marcher aprs
avoir t dcapit. Les canards
sans tte font de mme dans
la basse-cour. Dans un cadre
plus scientifique, des chats,
des souris ou des rats dont la
moelle pinire a t section-
ne peuvent recouvrer lusage
de leurs membres postrieurs,
quand bien mme la liaison
nerveuse avec le cerveau a t
interrompue.
Comment est-ce possible ? Il
y a un sicle, Charles Sherring-
ton [prix Nobel de mdecine
1932] avait mis en vidence la
persistance dune activit ryth-
mique dans la moelle pinire
sectionne. Des travaux plus
rcents ont montr que des
circuits, baptiss gnrateurs
de patrons locomoteurs (CPG),
situs dans la partie lombaire,
restent fonctionnels , explique
Grgoire Courtine (universit
de Zurich). Il est cosignataire
dun article dans la revue Na-
ture Neuroscience dcrivant la
faon dont des rats la moelle
pinire sectionne peuvent
marcher nouveau grce une
combinaison dinjections de
molcules pharmaceutiques,
de stimulations lectriques et
dentranement.
Dans le dispositif exprimen-
tal, les rats sont placs dans un
harnais qui les maintient en
positionverticale. Ils rpondent
en fait automatiquement la
stimulation quexerce sur leurs
pattes un petit tapis roulant.
Quand celui-ci fonctionne, les
pattes sadaptent immdiate-
ment audlement, sa vitesse
et son sens : elles peuvent
aussi bien marcher en avant
quen arrire. La moelle est
quasiment capabledecognition,
indique Grgoire Courtine : elle
peut utiliser les informations
sensorielles pour sadapter aux
conditions extrieures, sans
connexion avec le cerveau.
Ce phnomne nest pas indit.
Serge Rossignol (universit de
Montral), pionnier de ce type
de recherche, lavait mis en
vidence chez le chat ds les
annes 1980. Chez la souris et
le chat, la rcupration de cette
rythmicit fondamentale on
ne peut pas parler de marche
volontaire peut sobtenir sans
pharmacologie, grce un en-
tranement sur tapis roulant ,
rappelle-t-il. Cescircuits, prcise-
t-il, sont gntiquement dter-
mins : ils sont oprationnels
chez des chatons avant mme
que ceux-ci aient appris mar-
cher. Le mme phnomne
explique peut-tre la marche
automatique des nouveau-ns
humains.
D de la transposition
lhomme
EnFrance, lquipe de Didier Or-
sal (universit Pierre-et-Marie-
Curie, Paris) a mis au point une
technique de greffe de neurones
embryonnaires dans la moelle
pinire de rat. Ces neurones
mettent des prolongements et
sont capables de scrter de la
srotonine, un neurotransmet-
teur normalement produit par
le cerveau. Ce dispositif permet,
l aussi, dactiver le circuit lo-
comoteur du rat. Mais quand
leffet pharmacologique dispa-
rat, lanimal ne marche plus ,
prcise le chercheur.
Peut-ontransposer ces rsultats
chez lhomme ? Cest dj le
cas, en ce qui concerne len-
tranement propos certains
paraplgiques dont la moelle
nest que partiellement lse
pour recouvrer un peu de mo-
tricit. Ona aussi constat que
linjection de neurotransmet-
teurs permet de lutter contre les
escarres, apporte un meilleur
contrle des sphincters et rduit
latrophie musculaire , note
Didier Orsal.
Mais en ce qui concerne la r-
cupration de la locomotion,
le dfi reste immense : le cir-
cuit CPG napporte aucun
contrle volontaire du mouve-
ment. Il faudrait lasservir
des neuroprothses ca-
pables de faire le lien avec le
cerveau, dont le dveloppe-
ment reste embryonnaire. Une
autre approche vise la recon-
nexion des fibres nerveuses
lses, mais pour linstant une
protine baptise Nogo
empche ces repousses. Il ny
aura pas de miracle, prvient
Serge Rossignol, mais je suis
convaincu quon peut nourrir
des espoirs.
Herv Morin
(26 septembre 2009)
La moelle pinire,
gardienne de la locomotion
automatique
Lexcitation de circuits nerveux coups du cerveau permet des animaux de marcher
nouveau, sans volont propre.
POURQUOI
CET ARTICLE ?
Cet article montre limportance
de la moelle pinire, non pas
dans le contrle des rflexes,
mais dans la locomotion chez
les mammifres. En effet, lar-
ticle relate que chez des petits
mammifres, enabsencedetoute
connexion nerveuse entre la
moelle pinire et le cerveau, des
circuits neuronaux de la moelle
pinire restent fonctionnels et
sont lorigine de mouvements
de marche rythmiques, dits au-
tomatiques . Cette locomotion
automatiquencessitelaprsence
dune stimulation chimique ou
lectrique et saccrot avec len-
tranement. Ces observations
ouvrent des perspectives intres-
santes pour la rcupration de la
locomotionchezlhommeaprs
section de la moelle pinire.
Larticlefait lelienentreles fonc-
tions de la moelle pinire, tu-
dies lors dureexemyotatique,
et la motricit, tudie dans le
chapitre suivant.
L'ARTICLE DU
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
88
LESSENTIEL DU COURS
Neurone et bre musculaire : la communication nerveuse 89
LESSENTIEL DU COURS
Neurone et bre musculaire : la communication nerveuse
Motricit volontaire
et plasticit crbrale
De la volont au mouvement
Lenregistrement des potentiels daction au niveau
dun motoneurone montre que les circuits nerveux
impliqus dans le mouvement peuvent appartenir
larc rexe myotatique ou provenir de lencphale.
Les mouvements volontaires sont donc contrls par
lencphale. Lexploration du cortex crbral par les
techniques dimagerie mdicale permet didentier
les zones du cortex actives lors des mouvements
musculaires volontaires. Ltude de cas cliniques,
comme des lsions au niveau du cerveau entranant
des paralysies, ont galement particip la connais-
sanceducortexmoteur. Les mouvements volontaires
sont directement commands par laire motrice
primaire situe dans chaque hmisphre crbral.
Lacartographieprcisedecetteairemotriceprimaire
montre que chacune de ses rgions correspond
linnervation dune rgion prcise du corps. Plus la
motricit de cette rgion corporelle est complexe,
plus la rgion de laire motrice primaire qui lui est
ddie est large.
Les informations issues du cortex visuel situ lar-
rire du cerveau sont transmises laire prmotrice
et lairemotricesupplmentaire. Laireprmotrice,
implique plutt dans la rgulation de la posture,
dicte laire motrice la position optimale pour un
mouvement donn, tandis que laire motrice suppl-
mentaire inue sur la planicationet linitiationdes
mouvements en fonction des expriences passes.
Le cortex parital postrieur joue galement un
rle dans lexcution du mouvement volontaire, en
prenant en compte la position du corps, le geste
effectuer, etc., en intgrant les informations neuro-
sensorielles reues.
Les neurones pyramidauxde laire motrice primaire
projettent leurs axones vers le bulbe rachidien puis
vers la moelle pinire. Les voies motrices sont
croises : laire motrice primaire de lhmisphre
gauche contrle les mouvements de la partie droite
du corps et inversement. Les terminaisons synap-
tiques des neurones pyramidaux entrent en contact
au niveau de synapses avec les motoneurones. Au
niveau de la synapse, est libre une quantit de
neurotransmetteurs en adquation avec le train de
potentiels daction du neurone pr-synaptique. Le
motoneurone est ainsi en relation avec un grand
nombre de synapses certaines tant excitatrices,
dautre inhibitrices. Le motoneurone intgre alors
lensemble des informations nerveuses quil reoit
par un phnomne de sommation qui peut tre
spatialeoutemporelle. Lemessagenerveuxrsultant
au niveau du motoneurone est vhicul jusquau
muscle. Un mme motoneurone peut innerver plu-
A
lors que les rexes sont des mouvements inconscients,
nous ralisons des mouvements volontaires : saisir un
objet, marcher, courir, etc. Chez un tout jeune enfant, les
premiers gestes semblent trs maladroits : il ne sait pas attraper
une balle ni faire quelques pas. Mais, force de sentraner, de
faire des erreurs et de recommencer, il apprend matriser ses
gestes. Comment les mouvements volontaires sont-ils contr-
ls et comment expliquer lvolution des capacits motrices ?
Deglutition
Langue
Mchoire
Lvres
Visage
il
S
o
u
rc
il
C
o
u
P
o
u
c
e
D
o
ig
ts
M
a
in
P
o
ig
n
e
t
C
o
u
d
e
B
ra
s
p
a
u
le
Aire
motrice
primaire
Orteils
Cheville
G
e
n
o
u
H
a
n
c
h
e
T
ro
n
c
Cartographie de laire motrice primaire (Homunculus de
Peneld). Hmisphre gauche vue en coupe. Les parties du
corpslesplusmobiles, commelesmains, leslvresoulalangue
sont reprsentes hypertrophies.
Aire motrice
supplmentaire
Aire
prmotrice
Aire motrice
primaire
Cortex parital
postrieur
Planification
des mouvements
Organisationducortex moteur
sieurs bres ou cellules musculaires, mais une bre
musculaire nest toujours innerve que par un seul
motoneurone.
Plasticit et motricit crbrale
La comparaison des cartes motrices chez plu-
sieurs individus montre lexistence de diffrences
importantes. Pendant des priodes critiques (prin-
cipalement pendant lenfance), sous laction de
stimuli externes, des rorganisations des rseaux
neuronaux se produisent. Par exemple, lors de lap-
prentissage dun instrument comme le violon, les
violonistes droitiers utilisent les doigts de la main
gauche (notamment lannulaire et lauriculaire)
plus frquemment que les non-violonistes. Lors de
lactivation de lauriculaire gauche, le nombre de
dendrites actives auniveauducortexest plus lev
chez les violonistes (40 000 80 000 dendrites
actives) que chez les non-violonistes (1 000
35 000 dendrites actives). Lutilisation prf-
rentielle dun doigt augmente la reprsentation
corticale de la zone contrlant les mouvements
du doigt stimul, ainsi que lactivit des neurones
des zones concernes. Ainsi, le cortex moteur
prsente une plasticit crbrale, cest--dire la
capacit tablir de nouvelles connections entre
les neurones. De nouvelles synapses se mettent
en place : certaines vont disparatre, dautres vont
tre maintenues et renforces. De cette faon, les
diffrences dans le cortex moteur sacquirent au
cours du dveloppement, lors de lapprentissage
des gestes et de lentra-
nement. Larchitecture
du cerveau de chaque
individu rsultant de
son dveloppement est
donc unique. Cette plas-
ticit crbrale, que lon
retrouve dans dautres
parties du cerveau,
comme dans le cortex
visuel, est une proprit
gnrale du systme
nerveux central.
La plasticit crbrale
est aussi lorigine de
la capacit de rcup-
ration du cerveau aprs
un accident affectant
une petite partie du
cortex moteur et en-
tranant une perte fonc-
tionnelle. Par exemple,
un accident vasculaire
crbral (AVC) est un
trouble de lirrigation
sanguine du cerveau. Si
une partie du cerveau
cesse dtre irrigue
cause dun AVC, alors
les neurones meurent
et cette partie cesse de
fonctionner. Un AVC affectant une aire motrice
peut ainsi entraner une paralysie. Une rcupra-
tion du dcit moteur est parfois observe. La zone
affecte reste dtruite, mais des remaniements
affectant des zones voisines, permettent la rcu-
pration de la fonction motrice. Cette rcupration
est d'autant plus rapide qu'une reducationest faite
prcocement.
De manire gnrale, le nombre de neurones dimi-
nue avec l'ge (10 % environ de perte au cours de la
vie). Les apprentissages sont souvent plus aiss chez
les jeunes enfants que chez les adultes : il semble-
rait que les possibilits de plasticit crbrale di-
minuent avec lge, sans pour autant disparatre
totalement. Les neurones que nous possdons
constituent donc un vritable capital conserver
et entretenir.
Un motoneurone et ses connections
DEUXARTICLES DU MONDE
CONSULTER
Traiter plus tt Parkinson en stimulant le
cerveau p. 91
(Florence Rosier, 23 juin 2012)
Vers une police du cerveau p. 92
(Laurent Alexandre, 19 mai 2012)
ZOOM SUR
LES ACCIDENTS
VASCULAIRES
CRBRAUX (AVC)
Les accidents vasculaires c-
rbraux touchent environ
150 000 personnes en France
chaque anne : dans 80 % des
cas, lAVC est d une diminu-
tion de lapport sanguin (suite
un caillot par exemple) et dans
20 % des cas, lAVC est caus par
une hmorragie crbrale. LAVC
ne concerne pas uniquement
les personnes ges : plus de
10 000 personnes de moins de
45 ans sont victimes dun AVC
chaque anne.
Un des principaux facteurs de
risque est lhypertension ar-
trielle. LAVC prsente des si-
gnaux dalerte quil est parfois
possible de reconnatre (troubles
brutaux de la vision, de la parole,
de la marche, paralysie dune
partie du corps, etc.).
LAVC est une vritable urgence
mdicale : plus la victime est
prise en charge rapidement et
de manire adapte ds les tous
premiers signes, plus les chances
de survie et de limiter les s-
quelles sont leves.
LES HMIPLGIES
Un accident (AVC, traumatisme)
affectant le cortex moteur dun
seul hmisphre crbral en-
trane une paralysie du seul
ct du corps situ l'oppos de
lhmisphre crbral touch : il
sagit dune hmiplgie. En effet,
la commande des mouvements
volontaires est controlatrale
(le cortex moteur droit com-
mande les mouvements de la
partie gauche du corps) car les
voies motrices se croisent sous
le bulbe rachidien.
LES PARAPLGIES
Une lsion de la moelle pinire
peut tre lorigine de paralysie
qui affecte les parties gauche
et droite du corps. Ltendue
de la partie du corps paralyse
dpend de la hauteur laquelle
se situe la lsion. La paraplgie
consiste en la paralysie du bassin
et des jambes.
NOTIONS CLS
CORTEX MOTEUR,
AIRE MOTRICE
Partie du cortex crbral sp-
cialise dans la commande des
mouvements volontaires et qui
contient une aire motrice pri-
maire, une aire motrice suppl-
mentaire et une aire prmotrice.
CORTEX
Zone de faible paisseur, recou-
vrant la totalit du cerveau et
forme par la substance grise
(correspondant aux corps cellu-
laires des neurones). Lpaisseur
du cortex est denviron de 2
4 mm, sa supercie reprsente
2 200 m
2
, soit plus de 80 % de
la masse totale du cerveau. Le
cortex contient diffrentes aires,
spcialises dans des fonctions
spciques.
INTGRATION
NERVEUSE
Capacit des neurones laborer
un message nerveux rsultant
de la rception de plusieurs mes-
sages nerveux. On distingue la
sommation spatiale (prise en
compte des messages nerveux
issus de diffrentes synapses)
et la sommation temporelle
(prise en compte des diffrents
messages provenant dune mme
synapse sur un laps de temps
de quelques millisecondes). La
plupart du temps, la sommation
ralise est spatio-temporelle.
PLASTICIT CRBRALE
Capacit du cerveau voluer
dun point de vue anatomique et
fonctionnel sous leffet de lenvi-
ronnement, lors dun apprentis-
sage ou dune rducation. Cette
plasticit crbrale sexplique
par la rorganisation de circuits
neuronauxdus auxmodications
des synapses.
SYNAPSE
Zone de contact entre deux neu-
rones. Le message nerveux est
transmis dun neurone lautre
par librationpar le neurone pr-
synaptique dunneuromdiateur
qui agit sur le neurone postsy-
naptique.
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
90
UN SUJET PAS PAS
Neurone et bre musculaire : la communication nerveuse 91
LES ARTICLES DU
Neurone et bre musculaire : la communication nerveuse
1. Leneuronemoteurconduit unmessagenerveux:
a) cod en frquence de potentiel daction vers les
centres nerveux.
b) cod en amplitude de potentiel daction vers le
muscle effecteur.
c) prsentant toujours lammefrquenceet lamme
amplitude de potentiel daction.
d) cod en frquence de potentiel daction vers le
muscle effecteur.
2. Unebremusculaire:
a) reoit un message nerveux issu de plusieurs
motoneurones.
b) reoit un message nerveux issu dun seul moto-
neurone.
c) est capablederaliser uneintgrationdes messages
nerveux.
d) se relche lorsquelle reoit un message nerveux
dun motoneurone.
3. Auniveauducerveau, lescartesmotrices:
a) sont innes, prsentes ds la naissance.
b) restent identiques tout au long de la vie de lin-
dividu.
c) sont identiques cheztous les individus dunemme
espce.
d) peuvent voluer en fonction de lapprentissage.
4. Lecortexmoteur:
a) contient laire motrice primaire.
b) contient plusieurs aires qui communiquent entre
elles.
c) est situ en profondeur dans le cerveau.
d) est situ larrire du cerveau.
5. La partie ducorps la plus reprsente auniveaude
laire motrice primaire est :
a) le visage.
b) lpaule.
c) le genou.
d) lavant-bras.
6. Un accident vasculaire crbral dans le cortex
moteur de lhmisphre droit peut entraner :
a) une paralysie de la partie droite du corps.
b) une paralysie des deux membres suprieurs.
c) une paralysie des deux membres infrieurs.
d) une paralysie de la partie gauche du corps.
7. Une lsionde lamoelle pinire entrane :
a) une paralysie dun seul ct du corps.
b) une paralysie de tout le corps.
c) une paralysie de lapartie ducorps situe au-dessus
de la lsion.
d) uneparalysiedelapartieducorps situeendessous
de la lsion.
8. Laplasticit crbrale :
a) est due une augmentation du nombre de neu-
rones.
b) est due unremaniement des rseauxneuronaux.
c) sintensie avec lge.
d) est indpendante de lenvironnement.
Le corrig
1. d), 2. b), 3. d), 4. b), 5. a), 6. d), 7. d), 8. b)
Partie 1 : Motricit volontaire
et plasticit crbrale
Ce quil ne faut pas faire
Cocher deuxrponses oune cocher aucune rponse
Ne pas mobiliser les notions ducours.
AUTRES SUJETS POSSIBLES AU BAC
SUR CE THME
tude de documents
Cartographie du cortex moteur partir de don-
nes de limagerie crbrale.
Fonctionnement des synapses, intgration ner-
veuse.
Contrle dumuscle par les motoneurones.
Consquences des AVCet des lsions de la moelle
pinire sur la motricit.
Mise en vidence de la plasticit crbrale et vo-
lutionavec lge.
ZOOM SUR
LA MALADIE
DE PARKINSON
La maladie de Parkinson est une
maladie neurodgnrative
volution lente. Les principaux
symptmes sont les tremble-
ments de la main ou du pied,
la lenteur des mouvements et
la raideur du corps. La maladie
de Parkinson se caractrise par
une dgnrescence de neu-
rones situs la base du cer-
veau, contrlant les mouvements
automatiques et scrtant un
neurotransmetteur appel la
dopamine . Un traitement
base de L-dopa, un prcurseur de
la dopamine, amliore les symp-
tmes pendant plusieurs annes
puis perd de son efcacit. La
maladie de Parkinson concerne
principalement les personnes de
plus de 60 ans. Elle affecte 0,3 %
de la population gnrale. C'est
la deuxime maladie neurodg-
nrative la plus courante aprs la
maladie dAlzheimer.
LIRM
LIRM, ou imagerie par rson-
nance magntique , est une
technique dimagerie mdicale
permettant dobtenir des images
du cerveau, mais aussi dautres
organes. Sa rsolution est assez
leve : elle permet de voir des
structures dont la taille est in-
frieure un millimtre. Grace
lIRM, des coupes virtuelles du
cerveau peuvent tre ralises
pour caractriser des patholo-
gies. Limagerie par rsonnance
magntique fonctionnelle (IRMf)
permet de caractriser lactivit
des diffrentes parties du cer-
veau. Elle est donc particuli-
rement utilise en recherche
pour tudier le fonctionnement
crbral.
Dautres techniques dimageries
mdicales comme la TEP (tomo-
graphie par mission de posi-
tons) ou encore le scanner (ap-
pel aussi tomodensitomtrie
ou scanographie ) ont permis
ces trente dernires annes de
progresser dans les diagnostics
des atteintes crbrales et dans
les recherches en neurosciences.
Traiter plus tt Parkinson
en stimulant le cerveau
Une intervention prcoce pourrait faire gagner plusieurs annes de vie de qualit aux patients.
T
rs attendus des neuro-
logues, les rsultats de
ltude Earlystim ont t
rvls cette semaine Dublin
(Irlande), lors du 16
e
congrs
international sur la maladie
de Parkinson et les troubles du
mouvement. Prsents par le
professeur Gnther Deuschl,
de luniversit de Kiel (Alle-
magne), ils montrent lintrt
de traiter plus tt certains
malades parkinsoniens par la
technique de stimulation c-
rbrale profonde.
Cette intervention neurochi-
rurgicale consiste implanter
au sein mme du cerveau de
nes lectrodes qui dlivrent
un courant lectrique continu
jusque dans des structures c-
rbrales profondes, les noyaux
gris centraux. Lobjectif : modu-
ler, grce ce courant de haute
frquence, lactivit des circuits
neuronaux altrs par la ma-
ladie. Le tout premier patient
a t opr en 1993 par le pro-
fesseur Alim-Louis Benabid au
CHU de Grenoble. Aujourdhui,
quelque 400 parkinsoniens en
bncient chaque anne en
France.
Jusquici, ce traitement tait
rserv aux patients dont la
maladie tait trs volue,
un peu en dernier recours
aprs une longue souffrance,
tmoigne le docteur Michael
Schpbach, coordinateur du
volet franais de cette tude
lInstitut du cerveau et de la
moelle pinire (ICM, hpital
de la Piti-Salptrire, Paris).
Do notre interrogation :
ne pourrait-on pas oprer
les malades plus tt, quand
ils ont encore une vie sociale
et professionnelle ? Pour y
rpondre, ltude Earlystim a
inclus 251 malades en France et
en Allemagne.
En France, on compte 100 000
150000parkinsoniens. La sti-
mulationcrbrale ne concerne
quune petite fraction, envi-
ron 5 %, qui doivent rpondre
des critres dindication trs
stricts , prcise le professeur
Yves Agid, investigateur princi-
pal du volet franais de ltude
lICM. Rappelons que la mala-
die de Parkinson touche les cel-
lules dopaminergiques, celles
qui produisent un neurotrans-
metteur, la dopamine. On les
trouve dans la substance noire
du tronc crbral, une struc-
ture trs primitive prsente
chez tous les animaux. Ces cel-
lules forment linterrupteur
du comportement moteur ,
explique-t-il.
Ne peuvent tre oprs que
les patients qui nont que des
lsions dopaminergiques,
responsables des symptmes
typiques de la maladie len-
teur, rigidit et tremblement
de repos. Ils ne doivent avoir
ni troubles de la marche ou
de lquilibre ni troubles co-
gnitifs lis des lsions non
dopaminergiques. Ces patients
sont amliors de faon spec-
taculaire par la L-dopa, le trai-
tement substitutif qui pallie
le dficit de scrtion de la
dopamine.
Mais cette efcacit na quun
temps : aprs plusieurs an-
nes, des complications se
manifestent sous forme de
mouvements involontaires et
de uctuations motrices. Ha-
bituellement, les patients sont
oprs treize ans en moyenne
aprs le dbut de la maladie,
quand ces complications sont
dj svres , rapporte Michael
Schpbach. Dans Earlystim, ce
dlai a t rduit sept ans en
moyenne (parfois quatre ans),
avant que les effets indsirables
soient invalidants.
Trs bonne rponse
Les 251 patients de ltude ont
t rpartis par tirage au sort
en deux groupes : un groupe
sous traitement optimal par la
L-dopa seule et un groupe op-
r par stimulation crbrale,
suivie ou non dun traitement
par la L-dopa. Deux ans plus
tard, les deux groupes ont t
compars. Rsultat : alors que
la qualit de vie value par
les patients est maintenue
dans le groupe sous traitement
mdical seul, elle est amliore
de 26 % dans le groupe opr.
Pour la premire fois, les pa-
tients oprs tmoignent dune
amlioration de leur insertion
psychosociale , relve Michael
Schpbach.
Lvaluation mdicale montre
une trs bonne rponse la
stimulation prcoce sur les
symptmes parkinsoniens
comme sur les complications
de la L-DOPA. Mme un
stade prcoce de la maladie, il
peut tre bnque de prendre
le risque opratoire , estime
Michael Schpbach. Les pa-
tients concerns pourraient
gagner plusieurs annes de vie
de qualit , ajoute Yves Agid.
Pas question cependant dlar-
gir tout va les indications de
cette opration invasive. Son
succs reste conditionn deux
impratifs : le respect trs ri-
goureux des critres dindica-
tion et la prise en charge par
une quipe multidisciplinaire
trs exprimente.
Florence Rosier
(23 juin 2012)
POURQUOI
CET ARTICLE ?
LamaladiedeParkinsonest une
maladie qui se rvle vite han-
dicapante pour le patient, et la
prise de L-dopa, si elle amliore
les symptmes pendant uncer-
tain temps, ne les gurit pas et
cesse dagir aprs quelques an-
nes. Depuis une dizaine dan-
nes, il existeunethrapiebase
sur la stimulation lectrique
crbrale des structures pro-
fondes du cerveau, fortement
impliques dans le contrle de
la locomotion. Lactivit des
neurones dopaminergiques
impliqus est contrle par des
lectrodes implantes dans le
cerveau, permettant ainsi la
rgressiondes symptmes par-
kinsoniens. Mais cette thrapie
est pour linstant rserve des
patients fortement atteints par
la maladie et ne prsentant que
des troubles lis aux atteintes
des circuits dopaminergiques.
Cet article prsente les rsul-
tatsdunetudecliniquevisant
tendre la stimulation cr-
braledes patients prsentant
un stage plus prcoce de la
maladie. Les rsultats obtenus
montrent qulargir le champ
dapplication de cette thrapie
pourrait tre bnque ces
patients.
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
92
LES ARTICLES DU
LE GUIDE PRATIQUE
Neurone et bre musculaire : la communication nerveuse
Vers une police du cerveau
C
onstitudecent milliards
de neurones, le cerveau
tait un continent mys-
trieux. Lorigine de la pense et
de la conscience est reste une
nigme pour des gnrations de
scientiques. Ce nest qupartir
des annes 1980que Jean-Pierre
Changeux et Gerald Edelman
ont conceptualis une biologie
de la conscience. Les enjeux
sont immenses. Dune part,
comprendre comment sont
stockes nos motions et notre
mmoire est fondamental pour
lutter contre les maladies neu-
ropsychiatriques. Dautre part,
la technologie va permettre des
manipulations de nos cerveaux.
La comprhension du fonc-
tionnement crbral et la car-
tographie de lesprit humain
progressent au rythme de
laugmentation des capacits
informatiques. La stimulation
crbrale par implants qui
sont des puces lectroniques
implantes dans le cerveau
pour traiter les dpressions ou
les troubles nvrotiques graves
donne des rsultats trs encou-
rageants. Les ttraplgiques
peuvent commander un or-
dinateur ou une machine par
la pense, via un casque qui
analyse les ondes crbrales.
Rcemment, on a pu recons-
truire la pense grce au d-
codage des ondes enregistres
par des lectrodes crniennes.
La technologie ira au-del du
dcryptage des cerveaux : leur
manipulation semble sans li-
mite. Sa rgulation ne sera pas
consensuelle : on peut aussi
bien soutenir que le cerveau
doit rester un sanctuaire in-
violable que promouvoir des
techniques de neuro-renfor-
cement pour aider les enfants
moins favoriss. Dernirement,
lditorialiste de la prestigieuse
revue mdicale The Lancet sin-
quitait non des drives des
technologies de renforcement
crbral, mais des conditions
pour accorder aux tudiants
pauvres des bourses leur per-
mettant dy avoir accs !
Les neurotechnologies pour-
raient devenir une arme fatale
auservice dune ambitiontota-
litaire. Cest une menace indite
contre la libert : la police de
la pense sera technologique-
ment bientt prte. Lultime
frontire de la domination des
dictatures lesprit humain
serait pulvrise : on nose
imaginer ce que Staline, Mao,
Pol Pot ou Hitler auraient fait
des neurotechnologies. La pro-
tection de lintgrit crbrale
va devenir essentielle. Il fau-
dra encadrer les modications
mnsiques, mme lorsquelles
sont proposes au nomde lin-
trt des malades. Les militaires
travaillent aujourdhui sur les
techniques permettant de sup-
primer les souvenirs de guerre
traumatisants. Par ailleurs, la
socit ne se serait pas oppose
leffacement des souvenirs des
petites lles rescapes de laf-
faire Dutroux.
Et puis aprs ?
Aurait-il fallu si cela avait t
possiblesupprimer en1945 les
souvenirs atroces des rescaps
de la Shoah ? Pour le bien des
rares dports ayant survcu
peut-tre, mais pas pour lhu-
manit, dont lhistoireaurait t
falsie. Transformations bio-
logiques et lectroniques du
cerveau, ralit virtuelle, mani-
pulationdes souvenirs forment
un cocktail dtonant. La vrit
va devenir de plus en plus fra-
gile, et les erreurs que lon
trouvesur Wikipdiaparaissent
anodines auregarddes perspec-
tives des neurotechnologies.
Notre neuro-scurit, cest-
-direnotrelibert, deviendrale
cur des droits de lhomme de
lacivilisationbiotechnologique.
Maispourra-t-onfaireconance
ltat pour btir la neuro-
thique ? Autorisera-t-on, par
exemple, la justice lire dans
nos cerveaux ?
Laurent Alexandre
(19 mai 2012)
POURQUOI
CET ARTICLE ?
Cet article clt le thme des
neurosciences par une r-
flexion personnelle. Il pr-
sente la vision thique dun
scientifique face au progrs
spectaculaire de limagerie
crbrale de ces dernires
annes. Il est maintenant
possible davoir accs aux
structures du cerveau, de
connatre son fonctionne-
ment, de stimuler certaines
rgions prcises. L auteur
anticipe les possibilits tech-
niques venir et sinquite
du pouvoir que confre la
matrise de ces techniques et
les risques lis leur utilisa-
tion. Il appelle la cration
dune nouvelle branche de
la biothique : la neuro-
thique. Dans quelle mesure
a-t-on le droit davoir accs
lesprit dun tre humain ?
de le modifier ? Ce question-
nement est fondamental et
rappelle avec justesse que
toute innovation technolo-
gique doit saccompagner
dune rflexion thique sur
son utilisation et ses cons-
quences pour lindividu et
la socit.
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
94
LE GUIDE PRATIQUE
Le guide pratique 95
LE GUIDE PRATIQUE
Le guide pratique
preuve crite de SVT
Partie 1 : Matriser ses connaissances
Cette partie, qui porte sur une ou plusieurs parties
du programme spcique, a pour objectif dvaluer
les connaissances du candidat selon lune des mo-
dalits variables :
une question de synthse portant sur une ou
plusieurs parties du programme spcique ;
unQCMportant sur une partie ouplusieurs parties
duprogramme spcique, voire tout le programme.
Ce QCMpeut prendre appui sur des documents ;
lassociationdunQCM(avec ousans documents)
et dune question de synthse.
Dans la question de synthse sont values la ma-
trise des connaissances ducandidat et sa capacit
les mobiliser et les prsenter de manire cohrente
pour rpondre la questionpose. La qualit de la
rdaction est un critre important de lvaluation.
Il est donc essentiel didentier prcisment la
problmatique partir de lnonc pour dlimiter
le sujet et ainsi viter tout hors-sujet ou tout oubli
dune partie du sujet. La rponse attendue doit
contenir une introduction (qui dnit les termes
du sujet, pose clairement la problmatique et an-
nonce le plan), un dveloppement cohrent et une
conclusion(qui propose unbilanrpondant la pro-
blmatique puis une ouverture vers unthme voisin
du sujet tudi). Le dveloppement est structur en
diffrents paragraphes dont chacun dveloppe un
aspect dusujet de manire cohrente et argumente.
Des titres numrots mis en valeur peuvent tre
prsents mais ne sont pas exigibles. Le travail au
brouillon est impratif pour construire le plan de
la synthse : lisez bienlnonc, qui peut indiquer le
plan suivre, et mobilisez les notions vues encours
se rapportant au sujet. Prvoyez alors les schmas
ncessaires pour illustrer les notions choisies. Dans
tous les cas, suivez bien les consignes de lnonc
(par exemple un schma-bilan est souvent exig).
Les schmas doivent tre assez grands, clairs, en
couleurs et accompagns de lgendes et dun titre.
Dans le cas dunQCM, pour chaque question, quatre
propositions sont faites. Une seule proposition est
exacte, les autres sont fausses. Il ne faut jamais
cocher deux propositions car cela invalide auto-
matiquement la rponse. En gnral, seules les
bonnes rponses rapportent des points et labsence
de rponse ou une rponse errone nenlve pas
de point. Les candidats ont donc toujours intrt
cocher une proposition, mme sils ne sont pas
srs de leur rponse.
Partie 2 Exercice 1 : Raisonner
dans le cadre dun problme
scientique
Cet exercice de la partie 2 porte sur le programme
spcique et vise valuer la capacit du candidat
raisonner dans le cadre dun problme scien-
tique propos par le sujet, en sappuyant sur
lexploitation dun nombre rduit de documents.
La rponse peut se prsenter sous forme dun QCM
ou dune rponse rdige.
Dans le cas dune rponse rdige, il sagit de
construire unraisonnement rpondant auproblme
pos en exploitant les informations contenues
dans les documents. Il peut sagir par exemple de
comparer des lments, dargumenter, dextraire et
dorganiser des informations, de relier des informa-
tions entre elles partir des documents fournis, etc.
GRER
SON TEMPS
Consacrez 10 minutes en dbut
dpreuvelalecturedelensemble
du sujet et au choix de lordre
dans lequel vous allez traiter les
exercices. Fixez alors une dure
de travail pour chaque exercice et
veillez la respecter au mieux en
contrlant rgulirement lheure.
Traitezlensembledusujet mmesi
vousmatrisezmoinsbienunepar-
tie du programme. Avant la n de
lpreuve, prvoyez 5 10 minutes
pour relire votre copie et corriger
les fautes desens et dorthographe.
La veille de lpreuve, viter les
rvisions intensives tardives et es-
sayez darriver en forme, dtendu
mais motiv, le jour des preuves.
Prparez votre matriel pour com-
poser (y compris une boisson et
un petit en-cas). Avant le dbut de
lpreuve, prenezquelquesminutes
pour vous dtendre en respirant
calmement. Essayez de faire de
votre mieux quelles que soient les
difcults rencontres et protez
du temps imparti lpreuve pour
amliorer votre copie.
Prparation : 20minutes.
Prsentation : 20minutes.
Le candidat tire au sort un su-
jet de deux questions. Pour les
candidats qui nont pas choisi
la spcialit SVT, elles portent
sur deux parties diffrentes du
programme spcifique de SVT.
Pour les candidats ayant choisi la
spcialit SVT, une questionporte
sur le programme spcique et
lautre sur lun des thmes de
spcialit. Les sujets comportent
des documents semblables ceux
tudis en cours durant lanne.
Comme lpreuve a lieu dans une
salle comportant du matriel de
SVT, le candidat peut tre inter-
rog sur le matriel exprimental
et sonutilisation, sans tre amen
le manipuler.
GRER
SON STRESS
La qualit et la rigueur du raisonnement sont les
principaux critres de lvaluation de cet exercice.
Aucune restitution de connaissances nest exige
mais les connaissances du candidat peuvent tre
utiles pour analyser les documents. La rponse
attendue comporte, engnral, une courteintroduc-
tionposant la problmatique et undveloppement
structur en diffrents paragraphes (sans titres
apparents) qui prsentent le raisonnement.
Dans le cas dun QCM, il faut choisir partir de
lexploitation des documents la bonne rponse
parmi les quatre propositions prsentes pour
chaque question. Comme pour la partie 1, aucune
justication crite nest attendue, mais la lecture
attentive des documents et leur comprhension
sont indispensables pour slectionner les bonnes
rponses du QCM.
Partie 2 Exercice 2 : Pratiquer une
dmarche scientique
Cet exercice porte soit sur le programme spcique
pour les candidats nayant pas choisi la spcialit
SVT, soit sur un des thmes de spcialit pour les
candidats ayant choisi la spcialit SVT.
Lobjectif est dvaluer la capacit du candidat
pratiquer une dmarche scientique dans le cadre
de la rsolution dun problme scientique en
exploitant diffrents documents mis enrelationet
en mobilisant ses connaissances. Le candidat doit
exposer sa dmarche personnelle de rsolution du
problme enlaborant une vritable argumentation
qui conduit une rsolution possible du problme
pos et en proposant une conclusion rpondant
ce problme. Lnonc prcise souvent quaucune
tude exhaustive des documents nest attendue :
il ne sagit pas danalyser les documents dans leur
intgralit mais den extraire les informations
utiles la rsolution du problme pos, de les relier
entre elles et aux connaissances du candidat.
Dans cet exercice sont principalement values
la cohrence de la dmarche propose par le
candidat, sa capacit justier ses choix dans la
rsolution du problme et faire preuve desprit
critique. Plusieurs dmarches de rsolution du
problme peuvent tre possibles : le candidat
choisit celle qui lui parat la plus approprie en la
justiant rigoureusement laide des informations
extraites des documents.
La rponse attendue comporte, en gnral, une
introduction prsentant la problmatique, un d-
veloppement structur en diffrents paragraphes
(sans titres apparents) prsentant la dmarche
de rsolution, puis une conclusion rsumant la
rsolution du problme. Le candidat doit veiller
bien respecter la forme de la rponse exige par
lnonc : compte-rendu dune sortie sur le terrain,
analyse dun projet technique, etc. Les documents
ne sont pas analyser dans lordre donn par
lnonc mais selon lordre logique pour rsoudre
le problme. Untravail au brouillon est ncessaire
pour analyser les documents, slectionner les
informations issues des documents permettant
de traiter la problmatique, puis construire le
plan de la rponse laide des lments issus des
documents, mis en relation entre eux et avec les
lments de connaissances ncessaires.
Prparer lpreuve du bac
Au cours de lanne, au fur et mesure des cha-
pitres, ralisez des ches o gurent le plan du
cours, les notions essentielles, les dnitions des
mots cls ainsi que les schmas importants
connatre. Ce travail permet de slectionner les
notions essentielles, de les mmoriser et les revoir
rapidement lors des rvisions avant les preuves.
Retravaillez bien les corrections des valuations
faites en classe et entranez-vous sur des sujets de
type bac.
Dvelopper votre culture
gnrale
Les sujets font de plus en plus appel votre culture
gnrale. Celle-ci vous sera galement utile pour la
suite de vos tudes, pour des concours ou des en-
tretiens. Suivez les grandes lignes de lactualit
rgulirement (radio, tlvision, presse crite pa-
pier ou numrique) et approfondissez, par des
lectures, les sujets qui peuvent se rfrer au pro-
gramme. Les articles du Monde proposs ici visent
vous faciliter cette ouverture sur lactualit et la
mise relationentre avec les recherches scientiques
et le cours enseign.
LES SVT AU BAC
Coefficient : 6 ou 8 pour les
candidats ayant choisi la SVT
comme enseignement de sp-
cialit.
preuve pratique (valuation des
comptences exprimentales) :
sur 4 points ; dure : 1 h.
preuve crite : sur 16 points ;
dure : 3 h 30.
Note sur 8 points, elle value
la matrise des connaissances
acquises dans le cadre du pro-
gramme spcifique de SVT. Le
questionnement se prsente
sous forme dun QCM ou dune
question de synthse qui peu-
vent ventuellement sappuyer
sur un ou plusieurs documents.
Cette partie peut porter sur une,
plusieurs ou toutes les parties
du programme.
Note sur 8 points, elle value la
pratique du raisonnement scien-
tifique et de largumentation.
Elle comporte deux exercices :
le premier exercice, not sur
3 points, value la capacit du
candidat raisonner dans le
cadre dun problme scienti-
fique propos par le sujet, en
sappuyant sur lexploitation
dun nombre rduit de docu-
ments. Le questionnement se
prsente sous forme dun QCM
ou dune question ouverte ;
le second exercice, not sur
5 points, value la capacit du
candidat pratiquer une d-
marche scientifique dans le
cadre dun problme scienti-
fique partir de lexploitation
dun ensemble de documents et
en utilisant ses connaissances.
Pour les candidats qui nont
suivi que lenseignement spci-
fique de SVT, les deux exercices
de la partie 2 peuvent porter
sur la mme partie ou non du
programme spcifique. Pour les
candidats ayant choisi la sp-
cialit SVT, le second exercice
de la partie 2 porte sur lun des
thmes de spcialit.
PARTIE 1
DE LCRIT
PARTIE 2
DE LCRIT
PREUVE
ORALE DE
RATTRAPAGE
r
u
e
d
e
s
c
o
le
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
io
n
,
d
if
f
u
s
io
n
e
t
c
o
m
m
u
n
ic
a
t
io
n
s
t
r
ic
t
e
m
e
n
t
in
t
e
r
d
it
e
s
Crdits
GNTIQUE ET VOLUTION
Le brassage gntique et sa contribution la diversit gntique
p. 6 gauche, DR, droite Lzarts Cration ; p. 7, DR ; p. 8, DR ; p. 9, RDE.
Diversication gntique et diversication des tres vivants
p. 12 RDE ; p. 13 haut RDE, bas, DR, chimpanz Marcel Schauer/ Fotolia ;
p. 14 colonne Istockphoto/ Thinkstock, partie centrale Lzarts Cration ; p. 15 DR.
De la diversication des tres vivants lvolution de la biodiversit
p. 18 colonne haut RDE, colonne bas Istockphoto/ Thinkstock,partie centrale Lzarts Cration ;
p. 19 partie centrale RDE, colonne Istockphoto/ Thinkstockp. 20 DR ;
p. 21 partie centrale DR, colonne Istockphoto/ Thinkstock.
Un regard sur lvolution de lhomme
p. 24 RDE ; p. 25, DR ; p. 26 RDE ;
Les relations entre organisation et mode de vie, rsultat de lvolution :
lexemple de la vie xe chez les plantes
p. 30 DR ; p. 31 DR ; p. 32 DR ; p. 33 DR ;
LE DOMAINE CONTINENTAL ET SA DYNAMIQUE
La caractrisation du domaine continental :
lithosphre continentale, reliefs et paisseur crustale
p. 38 colonne et partie centrale droite Lzarts Cration, partie centrale gauche DR ; p. 39 partie centrale haut DR,
partie centrale bas et colonne RDE ; p.40 DR.
Contexte de la formation des chanes de montagnes et disparition des reliefs
p. 44 Lzarts Cration ; p. 45 partie centrale gauche Lzarts Cration, partie centrale droite DR, colonne RDE ;
p.46 carte
Lzarts Cration, autres visuels DR ; p.47 RDE
Le magmatisme en zone de subduction : uneproduction de nouveaux matriaux continentaux
p.50 partie centrale gauche DR, partie centrale droite RDE ; p. 51 Lzarts Cration ;
p. 52 en haut Lzarts Cration, en bas RDE
ENJEUX PLANTAIRES CONTEMPORAINS
Gothermie et proprits thermiques de la Terre
p. 56 DR ; p. 57 DR ; p. 58 gauche DR, droite RDE
La plante domestique
p. 60 Lzarts Cration ; p.61 gauche DR, droite RDE ; p. 62 RDE
LE MAINTIEN DE LINTGRIT DE LORGANISME :
QUELQUES ASPECTS DE LA RACTION IMMUNITAIRE
La raction inammatoire, un exemple de rponse inne
p. 66 haut et bas Hemera/ Thinkstock ; p. 67 RDE ; p. 69 DR
Limmunit adaptative, prolongement de limmunit inne
p. 72 DR ; p. 73 haut DR, bas Lzarts Cration ; p. 74 Istockphoto/ Thinkstock
Le phnotype immunitaire au cours de la vie
p. 79 haut DR, bas droite Lzarts Cration ; p. 80 RDE
NEURONE ET FIBRE MUSCULAIRE : LA COMMUNICATION NERVEUSE
Le rexe myotatique, un exemple de commande rexe du muscle
p. 84 Lzarts Cration ; p. 85 haut Lzarts Cration, bas DR ; p. 86 RDE
Motricit volontaire et plasticit crbrale
p. 88 RDE ; p. 89 RDE ; p. 90 Hemera/ Thinkstock
LE GUIDE PRATIQUE
p. 93 iStockphoto ; p. 94 Istockphoto/ Thinkstock ; p. 95 Purestock/ Thinkstock
r
u
e
d
e
s
c
o
l
e
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
i
o
n
,
d
i
f
f
u
s
i
o
n
e
t
c
o
m
m
u
n
i
c
a
t
i
o
n
s
t
r
i
c
t
e
m
e
n
t
i
n
t
e
r
d
i
t
e
s
r
u
e
d
e
s
c
o
l
e
s
&
L
e
M
o
n
d
e
,
2
0
1
3
.
R
e
p
r
o
d
u
c
t
i
o
n
,
d
i
f
f
u
s
i
o
n
e
t
c
o
m
m
u
n
i
c
a
t
i
o
n
s
t
r
i
c
t
e
m
e
n
t
i
n
t
e
r
d
i
t
e
s
T
erm
S
LESSENTIEL DU COURS
Des ches synthtiques
Les points et dnitions cls
Les repres importants
DES SUJETS DE BAC
Des questions types
Lanalyse des sujets
Les raisonnements
Les plans dtaills
Les piges viter
DES ARTICLES DU MONDE
Des articles du Monde
en texte intgral
Un accompagnement
pdagogique de chaque
article
UN GUIDE PRATIQUE
La mthodologie
des preuves
Astuces et conseils
H
O
R
S
-
S
R
I
E
H
o
r
s
-
s
r
i
e
L
e
M
o
n
d
e
,
a
v
r
i
l
2
0
1
3
Rviser son bac
avec
N
o
u
v
e
a
u
x
p
r
o
g
r
a
m
m
e
s
2
0
1
3
Sciences de la vie et
de la Terre
:
H
I
K
P
O
A
=
\
U
\
^
U
U
:
?
k
@
a
@
a
@
c
@
p
"
M
0
5
4
0
7
-
2
H
-
F
:
7
,
9
0
E
-
R
D
En partenariat avec
Vous aimerez peut-être aussi
- Philosophie PDFDocument114 pagesPhilosophie PDFfrancis de silva100% (2)
- Biochimie PDFDocument132 pagesBiochimie PDFAmanda Laleye58% (19)
- Reviser Son Bac Avec Le Monde PhilosophieDocument114 pagesReviser Son Bac Avec Le Monde PhilosophieGunnar Calvert100% (3)
- Ge OGRAPHIEDocument114 pagesGe OGRAPHIEfrancis de silva100% (2)
- Reviser Son Bac Avec Le Monde HISTOIRE PDFDocument98 pagesReviser Son Bac Avec Le Monde HISTOIRE PDFduthil_prof100% (4)
- Réviser Son Bac - PhilosophieDocument60 pagesRéviser Son Bac - PhilosophieRaja Rajan GPas encore d'évaluation
- 2nd - Cours SVT Lycée SecondeDocument232 pages2nd - Cours SVT Lycée SecondeWOGNIN HOFOUA AUBIN100% (4)
- HISTOIREDocument114 pagesHISTOIREfrancis de silva100% (1)
- Reviser Son Bac Avec Le Monde PHILOSOPHIE PDFDocument98 pagesReviser Son Bac Avec Le Monde PHILOSOPHIE PDFjujuPas encore d'évaluation
- REVISER Geo LE MONDEDocument98 pagesREVISER Geo LE MONDEeinguella einguellaPas encore d'évaluation
- 2016-05-01 Timbres de L 39 Orthographe PDFDocument84 pages2016-05-01 Timbres de L 39 Orthographe PDFFayed ElebiaPas encore d'évaluation
- Reviser Son Bac Avec Le Monde HISTOIREDocument0 pageReviser Son Bac Avec Le Monde HISTOIRErionetwo100% (3)
- FR Bac MondeDocument114 pagesFR Bac MondeMister MAAR Officielle100% (1)
- Culture Générale: - :HSMDOA UWUW (Document44 pagesCulture Générale: - :HSMDOA UWUW (Daw GuinePas encore d'évaluation
- Reviser Son Bac Avec Le Monde SCIENCES DE LA VIE-2 PDFDocument114 pagesReviser Son Bac Avec Le Monde SCIENCES DE LA VIE-2 PDFsebastien_houee100% (1)
- Tout Le Tronc Commun Tle Générale Johnny AnthonyDocument580 pagesTout Le Tronc Commun Tle Générale Johnny Anthonyguy tt100% (3)
- Reussir Au College PDFDocument10 pagesReussir Au College PDFLukPas encore d'évaluation
- Reviser Son Bac Avec Le Monde SCIENCES eCONOMIQUES ET SOCIALES TERMINALE PDFDocument98 pagesReviser Son Bac Avec Le Monde SCIENCES eCONOMIQUES ET SOCIALES TERMINALE PDFduthil_prof100% (1)
- Physique Chimie: Entraînement IntensifDocument544 pagesPhysique Chimie: Entraînement IntensifFranck MBELE100% (1)
- Reviser Son Bac Avec Le Monde SCIENCES eCONOMIQUES ET SOCIALES TERMINALE PDFDocument51 pagesReviser Son Bac Avec Le Monde SCIENCES eCONOMIQUES ET SOCIALES TERMINALE PDFibouziPas encore d'évaluation
- Reviser Son Bac Avec Le Monde PHILODocument98 pagesReviser Son Bac Avec Le Monde PHILOCéline Pornin100% (1)
- Reviser Son Bac Avec Le Monde HISTOIREDocument51 pagesReviser Son Bac Avec Le Monde HISTOIREGustavo100% (3)
- Manuel des difficultés de la langue française adapé au jeune âge et suivi d'un Recueil de locutions vicieusesD'EverandManuel des difficultés de la langue française adapé au jeune âge et suivi d'un Recueil de locutions vicieusesÉvaluation : 1 sur 5 étoiles1/5 (1)
- Reviser Son Bac Avec Le Monde GeOGRAPHIEDocument0 pageReviser Son Bac Avec Le Monde GeOGRAPHIErionetwo100% (4)
- Reviser Son Bac Avec Le Monde Sciences Economiques Et Sociales TerminaleDocument0 pageReviser Son Bac Avec Le Monde Sciences Economiques Et Sociales Terminalerionetwo100% (3)
- Reviser Bac SVTDocument51 pagesReviser Bac SVTNemo100% (3)
- (Réviser Son Bac Avec Le Monde) Rue Des Écoles-Mathématiques Terminale, Série S-Rue Des Écoles (2016)Document96 pages(Réviser Son Bac Avec Le Monde) Rue Des Écoles-Mathématiques Terminale, Série S-Rue Des Écoles (2016)hakima032Pas encore d'évaluation
- Reviser Son Bac Avec Le Monde GeOGRAPHIEDocument51 pagesReviser Son Bac Avec Le Monde GeOGRAPHIEiaidoiaido100% (2)
- ECS1 AnglaisDocument3 pagesECS1 AnglaisAnonymous XNijipPas encore d'évaluation
- Reviser Son Bac Avec Le Monde Sciences de La VieDocument114 pagesReviser Son Bac Avec Le Monde Sciences de La VieGunnar CalvertPas encore d'évaluation
- Francais - 1re - Le MondeDocument95 pagesFrancais - 1re - Le MondeVladimirPas encore d'évaluation
- Reviser Son Bac Avec Le Monde Philosophie PDFDocument98 pagesReviser Son Bac Avec Le Monde Philosophie PDFmokaPas encore d'évaluation
- Reviser Son Bac Avec Le Monde GeOGRAPHIE PDFDocument114 pagesReviser Son Bac Avec Le Monde GeOGRAPHIE PDFsebastien_houee100% (2)
- Le MondeDocument97 pagesLe MondeMartin RoxanaPas encore d'évaluation
- Révisions SVT Le Monde PDFDocument11 pagesRévisions SVT Le Monde PDFRahma HosniPas encore d'évaluation
- Reviser Son Bac Avec Le Monde FRANcAIS 1re ExtraitDocument18 pagesReviser Son Bac Avec Le Monde FRANcAIS 1re Extraitland987100% (2)
- Reviser Son Bac Avec Le Monde Sciences Economiques Et Sociales TerminaleDocument114 pagesReviser Son Bac Avec Le Monde Sciences Economiques Et Sociales TerminaleMaurice RogerPas encore d'évaluation
- Carte FRDocument59 pagesCarte FRAvram Simona100% (2)
- Romantisme: Les Grands Articles d'UniversalisD'EverandRomantisme: Les Grands Articles d'UniversalisPas encore d'évaluation
- Methodologie de La DissertationDocument6 pagesMethodologie de La DissertationMme et Mr LafonPas encore d'évaluation
- Reviser Son Bac Avec Le Monde Sciences Economiques Et Sociales TerminaleDocument51 pagesReviser Son Bac Avec Le Monde Sciences Economiques Et Sociales TerminaleOresb100% (1)
- Reviser Bac SVTDocument51 pagesReviser Bac SVTbob dylinPas encore d'évaluation
- SVT Lycee 2de Livretpdf PDFDocument8 pagesSVT Lycee 2de Livretpdf PDFMohamedaziz boutitiPas encore d'évaluation
- Oscar et la Dame rose d'Éric-Emmanuel Schmitt (Analyse approfondie): Approfondissez votre lecture de cette œuvre avec notre profil littéraire (résumé, fiche de lecture et axes de lecture)D'EverandOscar et la Dame rose d'Éric-Emmanuel Schmitt (Analyse approfondie): Approfondissez votre lecture de cette œuvre avec notre profil littéraire (résumé, fiche de lecture et axes de lecture)Pas encore d'évaluation
- Chagrin d'école de Daniel Pennac (Analyse de l'oeuvre): Analyse complète et résumé détaillé de l'oeuvreD'EverandChagrin d'école de Daniel Pennac (Analyse de l'oeuvre): Analyse complète et résumé détaillé de l'oeuvrePas encore d'évaluation
- Lizeaux - Baude Tle S Livre Du Professeur PDF - Télécharger, Lire TÉLÉCHARGER LIRE ENGLISH VERSION DOWNLOAD READ. DescriptionDocument8 pagesLizeaux - Baude Tle S Livre Du Professeur PDF - Télécharger, Lire TÉLÉCHARGER LIRE ENGLISH VERSION DOWNLOAD READ. DescriptionMon Compte0% (1)
- Enjeux de la place des savoirs dans les pratiques éducatives en contexte scolaire: Compréhension de l’acte d’enseignement et défis pour la formation professionnelle des enseignantsD'EverandEnjeux de la place des savoirs dans les pratiques éducatives en contexte scolaire: Compréhension de l’acte d’enseignement et défis pour la formation professionnelle des enseignantsPas encore d'évaluation
- TS Génétique SVTDocument23 pagesTS Génétique SVTDavid LemenagerPas encore d'évaluation
- Le dernier jour d'un condamné de Victor Hugo (Fiche de lecture): Analyse complète de l'oeuvreD'EverandLe dernier jour d'un condamné de Victor Hugo (Fiche de lecture): Analyse complète de l'oeuvrePas encore d'évaluation
- Chapitre II Etude de La Structure Des BactériesDocument10 pagesChapitre II Etude de La Structure Des BactériesAnthony MaryPas encore d'évaluation
- Document PDFDocument50 pagesDocument PDFsvonskPas encore d'évaluation
- Le Tour Du Monde en 80 JoursDocument6 pagesLe Tour Du Monde en 80 JoursMohaChebi0% (1)
- JustineDocument5 pagesJustineMourad TajPas encore d'évaluation
- TD Nomenclature Corrige 6Document2 pagesTD Nomenclature Corrige 6Aaouine AbderrazzakPas encore d'évaluation
- Rdii 31182Document2 pagesRdii 31182Mohammed TourarPas encore d'évaluation
- Manuel de Sciences de La Vie Et de La TerreDocument106 pagesManuel de Sciences de La Vie Et de La Terrejianastor100% (1)
- Entrer Dans Le Sup Apres Le Bac NPDCDocument124 pagesEntrer Dans Le Sup Apres Le Bac NPDCskarnskaPas encore d'évaluation
- Bac Blanc 1 Term ESDocument12 pagesBac Blanc 1 Term ESLe Détective sans MoustachePas encore d'évaluation
- Guide Général Des Enseignements de La Licence Histoire Du DPLSH À StClaude 2011-20122Document75 pagesGuide Général Des Enseignements de La Licence Histoire Du DPLSH À StClaude 2011-20122Université des Antilles100% (1)
- SVT 1ere S Chap1Document16 pagesSVT 1ere S Chap1maths726100% (3)
- Baby-sitter blues de Marie-Aude Murail (Fiche de lecture): Analyse complète de l'oeuvreD'EverandBaby-sitter blues de Marie-Aude Murail (Fiche de lecture): Analyse complète de l'oeuvrePas encore d'évaluation
- Ch1.1 La Mitose Cours PolyDocument2 pagesCh1.1 La Mitose Cours PolydadaPas encore d'évaluation
- La Levure BiologiqueDocument5 pagesLa Levure BiologiquePawan CoomarPas encore d'évaluation
- Série N°3Document3 pagesSérie N°3lmd2009Pas encore d'évaluation
- ReplicDocument12 pagesReplicKhalil BilalPas encore d'évaluation
- TD2 Genetique 2007Document5 pagesTD2 Genetique 2007Cia Mato75% (4)
- Cours 4 Génie GénétiqueDocument26 pagesCours 4 Génie GénétiqueAy IşığıPas encore d'évaluation
- Les PlasmidesDocument7 pagesLes PlasmidesDjerboua Taoufik100% (1)
- N6431972 PDF 1 - 1DM PDFDocument244 pagesN6431972 PDF 1 - 1DM PDFanon_736742916Pas encore d'évaluation
- Cours Transcription S5Document46 pagesCours Transcription S5ونزار عبد القادرPas encore d'évaluation
- Rouaissi Hajlaoui Mic. Hyg. Alim.08Document7 pagesRouaissi Hajlaoui Mic. Hyg. Alim.08hindPas encore d'évaluation
- TD Génie GénétiqueDocument4 pagesTD Génie GénétiqueMeryem ELkhalloufy100% (1)
- 2006CLF21636 PDFDocument318 pages2006CLF21636 PDFRoxana AtomeiPas encore d'évaluation
- SVTDocument6 pagesSVTHala AbdeldayemPas encore d'évaluation
- Valorisation Des DattesDocument171 pagesValorisation Des DattesجعدبندرهمPas encore d'évaluation
- Procaryotes Et EucaryotesDocument30 pagesProcaryotes Et EucaryotesHamza SerPas encore d'évaluation
- Etude de La Répartition Des Séquences AluDocument21 pagesEtude de La Répartition Des Séquences AluVonHammerPas encore d'évaluation
- Cours1 IntroductionDocument7 pagesCours1 IntroductionNibelMiaPas encore d'évaluation
- Chapitre 2 La RéplicationDocument18 pagesChapitre 2 La RéplicationHamza Ali100% (1)
- Bioremédiation PDFDocument31 pagesBioremédiation PDFmekaekPas encore d'évaluation
- Nom Anm OtcDocument122 pagesNom Anm OtcCristina RusnacPas encore d'évaluation
- Rush 00Document12 pagesRush 00kettuflyPas encore d'évaluation
- TS Fiche Révision Génétique Et ÉvolutionDocument5 pagesTS Fiche Révision Génétique Et ÉvolutionCarole Chamoun EbPas encore d'évaluation
- TS SVT Oblig Fiche Vocabulaire Partie 2Document4 pagesTS SVT Oblig Fiche Vocabulaire Partie 2tnerolf35Pas encore d'évaluation
- TableDocument6 pagesTabletomPas encore d'évaluation