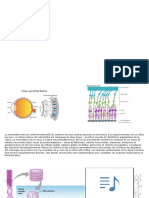
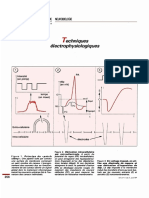
Vous aimerez peut-être aussi
- Utérus Artificiel: L'ère de l'utérus artificiel est sur le point et le monde de la matrice a commencéD'EverandUtérus Artificiel: L'ère de l'utérus artificiel est sur le point et le monde de la matrice a commencéPas encore d'évaluation
- 1 - Introduction À La GénomiqueDocument22 pages1 - Introduction À La Génomiquea.itatahine100% (1)
- TD Génomique Master UFR Sci Bio DR DagoDocument18 pagesTD Génomique Master UFR Sci Bio DR DagoPrince Otti's Officiel100% (1)
- Médecine Personnalisée: Utilisation du profil génétique pour le traitement de la maladieD'EverandMédecine Personnalisée: Utilisation du profil génétique pour le traitement de la maladiePas encore d'évaluation
- 1 - Introduction À La GénomiqueDocument39 pages1 - Introduction À La GénomiqueSamehPas encore d'évaluation
- GENOMIQUE - RésuméDocument5 pagesGENOMIQUE - RésuméRafa100% (1)
- Les Marqueurs MoléculairesDocument3 pagesLes Marqueurs MoléculairesAhmed Younsi100% (1)
- TD Biologie Moléculaire BPR 2021Document21 pagesTD Biologie Moléculaire BPR 2021rafaramalala1307Pas encore d'évaluation
- Exposé Application de Marqueurs Moléculaires Dans La Séléction Variétale Du RizDocument15 pagesExposé Application de Marqueurs Moléculaires Dans La Séléction Variétale Du RizJoOrdan AkàaPas encore d'évaluation
- FeuilletageDocument30 pagesFeuilletageSamir SoumanaPas encore d'évaluation
- ABM5. Genetique BacterienneDocument17 pagesABM5. Genetique BacterienneMichel Tatelos NdeukamPas encore d'évaluation
- Diagnostic Moléculaire - ArthurDocument133 pagesDiagnostic Moléculaire - ArthurBlaise MNPas encore d'évaluation
- La RT PCRDocument12 pagesLa RT PCRMbacké0% (1)
- $TD #1 D O.B PDFDocument2 pages$TD #1 D O.B PDFNacera CeraPas encore d'évaluation
- Gène Génétique BoucheffaDocument44 pagesGène Génétique Boucheffary yukPas encore d'évaluation
- Arbre Phylogenetique (1) - 4Document51 pagesArbre Phylogenetique (1) - 4bakheta hachemiPas encore d'évaluation
- CytogénétiqueDocument33 pagesCytogénétiqueFaiza HPas encore d'évaluation
- Exercice S 2016Document17 pagesExercice S 2016Aboubakar LawanePas encore d'évaluation
- Partie 1 - Biologie MoleculaireDocument48 pagesPartie 1 - Biologie MoleculaireZineb ZinebPas encore d'évaluation
- Replication ProcaryoteDocument5 pagesReplication ProcaryoteWissam TizaPas encore d'évaluation
- Biotech Mutagenese PDFDocument8 pagesBiotech Mutagenese PDFAghiles Ardj100% (2)
- Cours Clonage Et Banque de GènesDocument9 pagesCours Clonage Et Banque de GènesMadjidAvengersPas encore d'évaluation
- l3 BGV Cours 1 Bases Biologiques de La Culture in Vitro Vegetale - BousnaneDocument20 pagesl3 BGV Cours 1 Bases Biologiques de La Culture in Vitro Vegetale - Bousnanebelhout rauiyaPas encore d'évaluation
- 7 - PCRDocument23 pages7 - PCRVino DongaPas encore d'évaluation
- Marqueurs Moléculaires Pour L'amélioration de Blé TendreDocument27 pagesMarqueurs Moléculaires Pour L'amélioration de Blé Tendrefatima fek100% (1)
- Chapitre II La Réplication de LADNDocument37 pagesChapitre II La Réplication de LADNfekhar adlenPas encore d'évaluation
- Le Séquencage DADN 1Document4 pagesLe Séquencage DADN 1Kada BouameurPas encore d'évaluation
- AnémiesDocument18 pagesAnémiesmohamed essahyaoui100% (1)
- Série 03Document2 pagesSérie 03Moussaoui KhouiraPas encore d'évaluation
- Applications Des Techniques de Clonage Et de TransgenèseDocument17 pagesApplications Des Techniques de Clonage Et de Transgenèseelazyfy.douaee100% (1)
- Biologie Moleculaire: Cours deDocument98 pagesBiologie Moleculaire: Cours deMERYAM CHAWKIPas encore d'évaluation
- 2020-L1SV-Fascicule-Bio MolDocument8 pages2020-L1SV-Fascicule-Bio Molkirito 83Pas encore d'évaluation
- PCRDocument14 pagesPCRopenlabunistraPas encore d'évaluation
- Récepteurs Couplés Au Protéines G Partie 2Document7 pagesRécepteurs Couplés Au Protéines G Partie 2Fatima ZahraPas encore d'évaluation
- Lpro TD 1 Oct16 PDFDocument10 pagesLpro TD 1 Oct16 PDFAnonymous FYcmbifCtmPas encore d'évaluation
- Bioinformatique 3ème Licence Microbiologie 27 05 2018Document15 pagesBioinformatique 3ème Licence Microbiologie 27 05 2018BENYAHIA NESRINEPas encore d'évaluation
- Corrigé Exercice PCR 2020Document11 pagesCorrigé Exercice PCR 2020Lara TabetPas encore d'évaluation
- TD.02. BiomolDocument35 pagesTD.02. BiomolChawki Mokadem100% (1)
- ProgrammeDocument16 pagesProgrammeWalsy Sissoko100% (1)
- 5 MitochondrieDocument10 pages5 MitochondrieSõu ÂdPas encore d'évaluation
- Esp Réactives D'oxygène ROSDocument17 pagesEsp Réactives D'oxygène ROSmekaekPas encore d'évaluation
- Cours Biologie ApoptoseDocument11 pagesCours Biologie Apoptoseoussama bousseloubPas encore d'évaluation
- TP N°5 (1ère S)Document2 pagesTP N°5 (1ère S)sandra.le.du100% (2)
- Applications Du Génie GénétiqueDocument17 pagesApplications Du Génie GénétiqueSalma ZaPas encore d'évaluation
- Generalites BioinformatiqueDocument42 pagesGeneralites BioinformatiqueMariane LinganiPas encore d'évaluation
- Fiche TD 1Document4 pagesFiche TD 1nourzakiPas encore d'évaluation
- Cours de Biotechnologie Et Génie GénétiqueDocument29 pagesCours de Biotechnologie Et Génie GénétiqueMahaRahali100% (1)
- ChapIII - Le Fonctionnement Du Gène - Génétique - 1ère Année SB-1Document12 pagesChapIII - Le Fonctionnement Du Gène - Génétique - 1ère Année SB-1Amza ZinsonniPas encore d'évaluation
- Medecine-Intro Genetique MonohybridismeDocument23 pagesMedecine-Intro Genetique MonohybridismedzsergioPas encore d'évaluation
- Séquençage de l'ADN PDFDocument41 pagesSéquençage de l'ADN PDFChibani ZinebPas encore d'évaluation
- TD Les Transferts GénétiqueDocument4 pagesTD Les Transferts Génétiquescience علوم100% (1)
- Les Protozooses 2A Bio 2018Document31 pagesLes Protozooses 2A Bio 2018حمزة بن أحمدPas encore d'évaluation
- Epissage Chez Les EucaryotesDocument6 pagesEpissage Chez Les EucaryotesAlain Bertrand JuniorPas encore d'évaluation
- AFLP MarqueursDocument2 pagesAFLP MarqueursnourzakiPas encore d'évaluation
- Genetique1an HybridationDocument26 pagesGenetique1an HybridationHaniDjekrifPas encore d'évaluation
- !!!!!!!!cours Biologie Moleculaie S5Document46 pages!!!!!!!!cours Biologie Moleculaie S5Miro100% (1)
- TP 1mitoseMeioseFSAAM20Document6 pagesTP 1mitoseMeioseFSAAM20IMADPas encore d'évaluation
- C1 VirologieDocument18 pagesC1 VirologieSABRINA AYOUZPas encore d'évaluation
- Cours BM Master Les Techniques1Document29 pagesCours BM Master Les Techniques1Zineb IfriPas encore d'évaluation
- Compte RenduDocument4 pagesCompte RenduFANKELE ALASSANE KONEPas encore d'évaluation
- Cours Hemato M1BSS...Document21 pagesCours Hemato M1BSS...FANKELE ALASSANE KONEPas encore d'évaluation
- Expose PDFDocument7 pagesExpose PDFFANKELE ALASSANE KONEPas encore d'évaluation
- Technique Patch-ClampDocument2 pagesTechnique Patch-ClampFANKELE ALASSANE KONEPas encore d'évaluation
- TD Flavonoides Et TaninsDocument3 pagesTD Flavonoides Et TaninsFANKELE ALASSANE KONE100% (2)
- 2007 EveillardDocument160 pages2007 EveillardFANKELE ALASSANE KONEPas encore d'évaluation
- CM EPIDEMIO POP M1 SB UPGC - 2021 - vf01 - 19 05 2021Document208 pagesCM EPIDEMIO POP M1 SB UPGC - 2021 - vf01 - 19 05 2021FANKELE ALASSANE KONEPas encore d'évaluation
- Cours de Phytochimie 1 MASTERDocument22 pagesCours de Phytochimie 1 MASTERFANKELE ALASSANE KONE100% (1)
- Systeme SquelletteDocument44 pagesSysteme SquelletteFANKELE ALASSANE KONEPas encore d'évaluation
- TD - Système Nerveux - Sans TPDocument11 pagesTD - Système Nerveux - Sans TPFANKELE ALASSANE KONEPas encore d'évaluation
- Protocole ZafeleDocument19 pagesProtocole ZafeleFANKELE ALASSANE KONEPas encore d'évaluation
- 2182 Staphylococcus Aureus Resistants IsolesDocument69 pages2182 Staphylococcus Aureus Resistants IsolesFANKELE ALASSANE KONEPas encore d'évaluation
- COURS DEGRADATION ENVIRONNEMENT 6emeDocument4 pagesCOURS DEGRADATION ENVIRONNEMENT 6emeFANKELE ALASSANE KONEPas encore d'évaluation
- StaureusDocument5 pagesStaureusFANKELE ALASSANE KONEPas encore d'évaluation
- MR 258 Super5 4 5 9Document259 pagesMR 258 Super5 4 5 9Van Aster EricPas encore d'évaluation
- Justificatif LienDocument1 pageJustificatif Lienimmavillena0Pas encore d'évaluation
- Microscope Électronique À BalayageDocument29 pagesMicroscope Électronique À BalayageS0UM0HPas encore d'évaluation
- La très petite entreprise (TPE) en AfriqueDocument8 pagesLa très petite entreprise (TPE) en Afriquetheodoreahoton1Pas encore d'évaluation
- Facture Freemobile 20231230Document2 pagesFacture Freemobile 20231230ashwineuPas encore d'évaluation
- Liste Des Variétés de PoiresDocument3 pagesListe Des Variétés de Poiresmohamedadjeb2001Pas encore d'évaluation
- Heyman - Measurement of The Influence of Lateral Earth PressureDocument5 pagesHeyman - Measurement of The Influence of Lateral Earth PressureMarcelaMarquesPas encore d'évaluation
- Matrice Multi CritèresDocument57 pagesMatrice Multi CritèresOthmane AouinatouPas encore d'évaluation
- La Grammaire Des Premiers Temps V1 Reponses Corriges PDFDocument32 pagesLa Grammaire Des Premiers Temps V1 Reponses Corriges PDFadastraperalasporci100% (2)
- Expose Equipements de Production Tarek SenigraDocument5 pagesExpose Equipements de Production Tarek Senigratarek senigraPas encore d'évaluation
- Sicareme Assurances: Certificat D'Assurances Etudes + Stages POLICE #470.2020.00000049Document1 pageSicareme Assurances: Certificat D'Assurances Etudes + Stages POLICE #470.2020.00000049Aladine MardyPas encore d'évaluation
- Techniques D Obfuscation de Code Chiffrer Du Clair Avec Du ClairDocument20 pagesTechniques D Obfuscation de Code Chiffrer Du Clair Avec Du ClairMouhamed Rassoul GueyePas encore d'évaluation
- Techniques D'elevage Intensif Et D'alimentation de Poissons Et de CrustacesDocument19 pagesTechniques D'elevage Intensif Et D'alimentation de Poissons Et de CrustacesDao SouleymanePas encore d'évaluation
- Cours n2Document4 pagesCours n2Sophie BarrierePas encore d'évaluation
- Chap2 Presentation Du CloudDocument16 pagesChap2 Presentation Du CloudImene Ben SalemPas encore d'évaluation
- Exposé SVT Les Minerais A MadagascarDocument7 pagesExposé SVT Les Minerais A MadagascarJohan RaharisonPas encore d'évaluation
- Inf302 TDDocument34 pagesInf302 TDRio LoboPas encore d'évaluation
- 0709 ArduinoDocument6 pages0709 Arduinohamza ayechePas encore d'évaluation
- Economie IndustrielleDocument4 pagesEconomie IndustrielleLatifa HadekPas encore d'évaluation
- Rituel CBCS Pour Chevaliers Et EcuyersDocument94 pagesRituel CBCS Pour Chevaliers Et EcuyerskimonthPas encore d'évaluation
- Notice de Sécurité IncendieDocument114 pagesNotice de Sécurité IncendieYasser MeftouhPas encore d'évaluation
- Les Figures de StyleDocument4 pagesLes Figures de StylemouadhPas encore d'évaluation
- Corrigé UE11 - DCG 2011Document17 pagesCorrigé UE11 - DCG 2011Med Reda BouasriaPas encore d'évaluation
- Blondel - La Religion. Le Problème de La MystiqueDocument40 pagesBlondel - La Religion. Le Problème de La MystiquemolinerPas encore d'évaluation
- TD1 Me ÜmoiresDocument3 pagesTD1 Me ÜmoiresalaeelhassanyPas encore d'évaluation
- Parkside PKS2000-4 (Tronçonneuse)Document20 pagesParkside PKS2000-4 (Tronçonneuse)Georges KouroussisPas encore d'évaluation
- 201 Coute - NR 4 2023Document68 pages201 Coute - NR 4 2023luPas encore d'évaluation
- 5analyse TypomorphologiqueDocument10 pages5analyse Typomorphologiqueyasmine bouheloufPas encore d'évaluation
- Voie LactéeDocument34 pagesVoie Lactéejulian perezPas encore d'évaluation
- Projet 02. Séq 02. BOUSEKKINE Anfel 2Document25 pagesProjet 02. Séq 02. BOUSEKKINE Anfel 2Renda ReziguiPas encore d'évaluation
- Améliorer votre mémoire: Un Guide pour l'augmentation de la puissance du cerveau, utilisant des techniques et méthodesD'EverandAméliorer votre mémoire: Un Guide pour l'augmentation de la puissance du cerveau, utilisant des techniques et méthodesÉvaluation : 5 sur 5 étoiles5/5 (2)
- Cahier de jeux de stimulation cognitive: Sujets Alzheimer, désorientés, démences, amnésiesD'EverandCahier de jeux de stimulation cognitive: Sujets Alzheimer, désorientés, démences, amnésiesPas encore d'évaluation
- Les 10 Secrets pour une Vie Plus Heureuse avec la Maladie de ParkinsonD'EverandLes 10 Secrets pour une Vie Plus Heureuse avec la Maladie de ParkinsonPas encore d'évaluation
- Manuel pour les débutants Fabriquez des savons naturelsD'EverandManuel pour les débutants Fabriquez des savons naturelsÉvaluation : 3 sur 5 étoiles3/5 (2)
- La thérapie libre: La Thérapie de LIBération et Résilience EmotionnelleD'EverandLa thérapie libre: La Thérapie de LIBération et Résilience EmotionnelleÉvaluation : 5 sur 5 étoiles5/5 (1)
- Histoire de la psychologie scientifique: De la naissance de la psychologie à la neuropsychologie et aux champs d'application les plus actuelsD'EverandHistoire de la psychologie scientifique: De la naissance de la psychologie à la neuropsychologie et aux champs d'application les plus actuelsPas encore d'évaluation
- La vie des abeilles: Prix Nobel de littératureD'EverandLa vie des abeilles: Prix Nobel de littératureÉvaluation : 4 sur 5 étoiles4/5 (41)
- Nutrition Sportive Fondamentaux et guide pratique du succèsD'EverandNutrition Sportive Fondamentaux et guide pratique du succèsÉvaluation : 5 sur 5 étoiles5/5 (1)
- Guide De Démarrage De L'Apiculture: Le Guide Complet Pour Élever Des AbeillesD'EverandGuide De Démarrage De L'Apiculture: Le Guide Complet Pour Élever Des AbeillesPas encore d'évaluation
- Harmonisation Energétique des Lieux: Habitat et haut-lieux sacrés 2020D'EverandHarmonisation Energétique des Lieux: Habitat et haut-lieux sacrés 2020Évaluation : 2.5 sur 5 étoiles2.5/5 (3)
- Géologie de l'Amérique: Les Grands Articles d'UniversalisD'EverandGéologie de l'Amérique: Les Grands Articles d'UniversalisPas encore d'évaluation
- Microbiologie médicale I: agents pathogènes et microbiome humainD'EverandMicrobiologie médicale I: agents pathogènes et microbiome humainÉvaluation : 5 sur 5 étoiles5/5 (1)
- Pourquoi suis-je différent?: Accepter et découvrir la puissance de l'hypersensibilitéD'EverandPourquoi suis-je différent?: Accepter et découvrir la puissance de l'hypersensibilitéPas encore d'évaluation
- Les Graines d'Abricot - Remède de Cancer avec de la Vitamine B17 ?: La Médecine Antique Que l'Industrie Pharmaceutique Moderne CacheD'EverandLes Graines d'Abricot - Remède de Cancer avec de la Vitamine B17 ?: La Médecine Antique Que l'Industrie Pharmaceutique Moderne CachePas encore d'évaluation
- Le CODE DE DIEU: Le secret de notre passé, la promesse de notre avenirD'EverandLe CODE DE DIEU: Le secret de notre passé, la promesse de notre avenirÉvaluation : 4 sur 5 étoiles4/5 (23)
- Qu'arrive t'il a la terre arable ?: Le problème de l'érosion du sol.D'EverandQu'arrive t'il a la terre arable ?: Le problème de l'érosion du sol.Évaluation : 5 sur 5 étoiles5/5 (1)
- La Conscience Et L'Univers Existent Sans Commencement Ni FinD'EverandLa Conscience Et L'Univers Existent Sans Commencement Ni FinPas encore d'évaluation
- Encyclopédie de la magie des cristaux, des pierres précieuses et des métauxD'EverandEncyclopédie de la magie des cristaux, des pierres précieuses et des métauxÉvaluation : 4 sur 5 étoiles4/5 (116)
- Transition énergétique ET Changement climatique: Développements et Perspectives d'AvenirD'EverandTransition énergétique ET Changement climatique: Développements et Perspectives d'AvenirPas encore d'évaluation