Vous aimerez peut-être aussi
- (Chapitre 2 Organisation Fonctionnelle Des Systèmes Reproducteurs) PDFDocument14 pages(Chapitre 2 Organisation Fonctionnelle Des Systèmes Reproducteurs) PDFSamra LadjelPas encore d'évaluation
- 5-Physiologie Des GonadesDocument17 pages5-Physiologie Des GonadesNateche NassiraPas encore d'évaluation
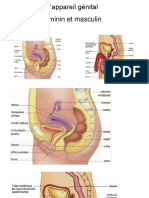
- Chap 2 Appareil Génital FémininDocument10 pagesChap 2 Appareil Génital Fémininabdoul djalil soumanaPas encore d'évaluation
- Appareil ReproducteurDocument9 pagesAppareil ReproducteurAntonella GiulianiPas encore d'évaluation
- L - Ovaire - 2019 2020Document12 pagesL - Ovaire - 2019 2020aacer3976Pas encore d'évaluation
- 10 - Ovogenèse Et FolliculogénèseDocument18 pages10 - Ovogenèse Et Folliculogénèsedjamila benaissaPas encore d'évaluation
- Histologie de l’Appareil (2)Document33 pagesHistologie de l’Appareil (2)qty9jgkpnzPas encore d'évaluation
- LA REPRODUCTIONDocument19 pagesLA REPRODUCTIONDiogob DiopPas encore d'évaluation
- MORPHOLOGIE FONCTIONNELLE DES ORGANES DU SYSTÈME GENITAL FÉMININDocument17 pagesMORPHOLOGIE FONCTIONNELLE DES ORGANES DU SYSTÈME GENITAL FÉMININanouarPas encore d'évaluation
- ReproductionDocument55 pagesReproductionNoura Azaouag100% (1)
- SYSTEME REPRODUCTEUR Bon - 102716Document13 pagesSYSTEME REPRODUCTEUR Bon - 102716Jude AgbodjiPas encore d'évaluation
- Ovogenese Et Folliculogenese FinaleDocument62 pagesOvogenese Et Folliculogenese Finalemaryam.gynoPas encore d'évaluation
- Nouveau Document Microsoft WordDocument21 pagesNouveau Document Microsoft WordDoua SouayahPas encore d'évaluation
- 15.03.a.cycle OvarienDocument29 pages15.03.a.cycle OvarienMamadou SyPas encore d'évaluation
- Appareil Genital de La Femme 2024 - 1Document8 pagesAppareil Genital de La Femme 2024 - 1KobePas encore d'évaluation
- Partie 2 OstetriqueDocument59 pagesPartie 2 Ostetriquebeatricekasongo465Pas encore d'évaluation
- 4 - Ovogenese Cours 2023Document3 pages4 - Ovogenese Cours 2023Amira AmirPas encore d'évaluation
- 2 - App Genital Fem en 10 PageDocument10 pages2 - App Genital Fem en 10 PageYo miPas encore d'évaluation
- 3a p2 Ue7 Cours 12 Physiologie en Gynécologie Ronéo PDFDocument14 pages3a p2 Ue7 Cours 12 Physiologie en Gynécologie Ronéo PDFAbaidi Sihem2108Pas encore d'évaluation
- OvogeneseDocument4 pagesOvogenesedyhp2pswsrPas encore d'évaluation
- Embryo23 04-Ovulation Cycle MenstruelDocument5 pagesEmbryo23 04-Ovulation Cycle MenstruelChouaib MeraoumiaPas encore d'évaluation
- Chapitre II OvogenèseDocument4 pagesChapitre II OvogenèseDel HadPas encore d'évaluation
- 3 APPAREIL GENITAL FEMININ WordDocument13 pages3 APPAREIL GENITAL FEMININ WordFaïsole Adimou Ola100% (2)
- 3 Appareil Genital Feminin WordDocument13 pages3 Appareil Genital Feminin WordBourama KoumarePas encore d'évaluation
- Apparei L Genital Misa JourDocument21 pagesApparei L Genital Misa JourBakary SanogoPas encore d'évaluation
- Resumé Reproduction Par AnouarDocument5 pagesResumé Reproduction Par Anouaran_angel_from_the_hell100% (1)
- 2 Fonction Génitale FemelleDocument15 pages2 Fonction Génitale Femellegeorge.guergourPas encore d'évaluation
- Cours D'obstetrique DiapoDocument170 pagesCours D'obstetrique Diapojuniorebinda93% (14)
- Histologie de L'ovaireDocument25 pagesHistologie de L'ovairemalakzerizer2005Pas encore d'évaluation
- Embryologie Générale 2Document49 pagesEmbryologie Générale 2Sofiane BousifePas encore d'évaluation
- ReproductionDocument14 pagesReproductionjamalPas encore d'évaluation
- Chap 5Document14 pagesChap 5Mamadou lamine DiattaPas encore d'évaluation
- SVT Ghediri - Bac Sciences Reproduction Humaine Chez La Femme-ConvertiDocument10 pagesSVT Ghediri - Bac Sciences Reproduction Humaine Chez La Femme-ConvertiRihane FarahPas encore d'évaluation
- Retenir Seq 5 Part 3Document6 pagesRetenir Seq 5 Part 3Julia NicolettiPas encore d'évaluation
- La Reproduction de L'homme Et L'éducation À La Sexualité CM2Document18 pagesLa Reproduction de L'homme Et L'éducation À La Sexualité CM2lezardkingPas encore d'évaluation
- Physiologie de La FonctionDocument12 pagesPhysiologie de La FonctionhicamPas encore d'évaluation
- 5 OvulationDocument6 pages5 OvulationAkram ChetouanePas encore d'évaluation
- Embryo23 03-GametogeneseDocument9 pagesEmbryo23 03-GametogeneseChouaib MeraoumiaPas encore d'évaluation
- Femme2 Adoc PDFDocument5 pagesFemme2 Adoc PDFsonilolaPas encore d'évaluation
- Biologie - La Reproduction - Les cycles sexuels et la régulation de la sécrétion des hormones sexuelles_1600814336114Document7 pagesBiologie - La Reproduction - Les cycles sexuels et la régulation de la sécrétion des hormones sexuelles_1600814336114ousmanekangama2Pas encore d'évaluation
- Cycle MenstruelDocument76 pagesCycle MenstruelMahamat Barkadei HangataPas encore d'évaluation
- 1.Fécondation nidation placentationDocument7 pages1.Fécondation nidation placentationAmihaf AmihafPas encore d'évaluation
- Anat Obst App GenDocument15 pagesAnat Obst App Gensalpro2Pas encore d'évaluation
- Assistance Médicale À La ProcréationDocument2 pagesAssistance Médicale À La Procréationamelamrani1918Pas encore d'évaluation
- Obstetrique 2015, Pr. Elongi PDFDocument242 pagesObstetrique 2015, Pr. Elongi PDFBienfait Kabwika100% (5)
- Cours Folliculogénèse-Lovulation-CJ-Fécondation-2011. Actualisé en Janvier 2020-Lecture SeuleDocument11 pagesCours Folliculogénèse-Lovulation-CJ-Fécondation-2011. Actualisé en Janvier 2020-Lecture SeuleMSDPas encore d'évaluation
- Fecondation Et EmbriologieDocument22 pagesFecondation Et EmbriologieDjily MbayePas encore d'évaluation
- Fichier Produit 2176Document26 pagesFichier Produit 2176falida compaorePas encore d'évaluation
- TP BIOS 3096_Séance 2_2022Document13 pagesTP BIOS 3096_Séance 2_2022Juan BlackspiritPas encore d'évaluation
- Fonction Ovarienne Physiologie Et ExplorationDocument12 pagesFonction Ovarienne Physiologie Et ExplorationAlina Topor Psi-MedicalPas encore d'évaluation
- Cours SVTDocument8 pagesCours SVTKukukovaNicolasPas encore d'évaluation
- 3397110Document25 pages3397110Nabil holmesPas encore d'évaluation
- Syst ReprodDocument28 pagesSyst ReprodGregory GebertPas encore d'évaluation
- Developpement Embryonnaire Chez L'humain Nouvelle VersionDocument55 pagesDeveloppement Embryonnaire Chez L'humain Nouvelle VersionMeguerouni Ghania100% (1)
- Ovogénèse Cours WordDocument15 pagesOvogénèse Cours WordNadjib A. NedjarPas encore d'évaluation
- Capture d’écran . 2024-02-12 à 23.51.59Document31 pagesCapture d’écran . 2024-02-12 à 23.51.59imanebouhachmoud157Pas encore d'évaluation
- Embryo23 05-2mois Devellopement PDFDocument30 pagesEmbryo23 05-2mois Devellopement PDFChouaib MeraoumiaPas encore d'évaluation
- Procréation: Les Grands Articles d'UniversalisD'EverandProcréation: Les Grands Articles d'UniversalisPas encore d'évaluation
- Sexualisation en biologie: Les Grands Articles d'UniversalisD'EverandSexualisation en biologie: Les Grands Articles d'UniversalisPas encore d'évaluation
- Appareil génital: Les Grands Articles d'UniversalisD'EverandAppareil génital: Les Grands Articles d'UniversalisPas encore d'évaluation
- Physiologie de La Glande ThyroïdeDocument5 pagesPhysiologie de La Glande ThyroïdeZied Bouabene100% (1)
- Clairances GlomérulairesDocument1 pageClairances GlomérulairesZied BouabenePas encore d'évaluation
- La SomesthésieDocument3 pagesLa SomesthésieZied Bouabene100% (1)
- Réabsorption Rénale Du GlucoseDocument1 pageRéabsorption Rénale Du GlucoseSaken667Pas encore d'évaluation
- Généralités Sur La Physiologie RénaleDocument3 pagesGénéralités Sur La Physiologie RénaleZied BouabenePas encore d'évaluation
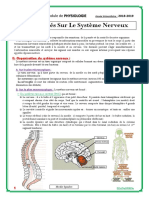
- Généralités Sur Le Système NerveuxDocument3 pagesGénéralités Sur Le Système NerveuxSaken667Pas encore d'évaluation
- Clairance Osmolaire Et Clairance de L'Eau LibreDocument1 pageClairance Osmolaire Et Clairance de L'Eau LibreZied Bouabene67% (3)
- Les Noyaux Gris CentrauxDocument5 pagesLes Noyaux Gris CentrauxZied BouabenePas encore d'évaluation
- Mécanisme D'Acidification Des UrinesDocument3 pagesMécanisme D'Acidification Des UrinesZied Bouabene100% (1)
- Physiologie Des Récepteurs SensorielsDocument2 pagesPhysiologie Des Récepteurs SensorielsSaken667Pas encore d'évaluation
- Excrétion Urinaire Du Potassium PDFDocument2 pagesExcrétion Urinaire Du Potassium PDFZied BouabenePas encore d'évaluation
- Physiologie de La SurrénaleDocument7 pagesPhysiologie de La SurrénaleSaken667100% (1)
- Circulation RénaleDocument3 pagesCirculation RénaleZied BouabenePas encore d'évaluation
- Physiologie Du Complexe Hypothalamo-HypophysaireDocument5 pagesPhysiologie Du Complexe Hypothalamo-HypophysaireZied Bouabene100% (1)
- Filtration GlomérulaireDocument3 pagesFiltration GlomérulaireZied BouabenePas encore d'évaluation
- La MictionDocument2 pagesLa MictionZied Bouabene100% (1)
- Réabsorption Rénale Du Calcium Et Des PhosphatesDocument2 pagesRéabsorption Rénale Du Calcium Et Des PhosphatesZied BouabenePas encore d'évaluation
- Microzymas Dans Leurs Rapports Avec L'heteroghologie .., Les - Bechamp, Antoine, 1816-1908Document1 075 pagesMicrozymas Dans Leurs Rapports Avec L'heteroghologie .., Les - Bechamp, Antoine, 1816-1908jncvnPas encore d'évaluation
- 4 - La Réparation de l'ADNDocument43 pages4 - La Réparation de l'ADNrajaa100% (2)
- l2 s3 Ecolgen Amsellem IntroDocument54 pagesl2 s3 Ecolgen Amsellem IntroNabil holmesPas encore d'évaluation
- 100 QCM Corrigés ImmunologieDocument17 pages100 QCM Corrigés Immunologiechaima kouchPas encore d'évaluation
- Skuldtech Manuel Utilisation DAVE 310811Document24 pagesSkuldtech Manuel Utilisation DAVE 310811samir801Pas encore d'évaluation
- CMurogénitalDocument11 pagesCMurogénitalflora.batailleaoustinPas encore d'évaluation
- Plantes Menacées Du QuébecDocument184 pagesPlantes Menacées Du Québecninam1Pas encore d'évaluation
- Le Processus de Cancerisation PDFDocument161 pagesLe Processus de Cancerisation PDFkarolPas encore d'évaluation
- Corrige Devoir 5 28Document4 pagesCorrige Devoir 5 28niyomo7937Pas encore d'évaluation
- Chap 1generalites Sur Les Champignons Mycologie MedicaleDocument16 pagesChap 1generalites Sur Les Champignons Mycologie MedicaleNgameni LeonelPas encore d'évaluation
- YOUCEFDocument113 pagesYOUCEFYoucef abPas encore d'évaluation
- Exercice 1Document2 pagesExercice 1Amadou CISSEPas encore d'évaluation
- Activité 1 Remobilisation Des AcquisDocument4 pagesActivité 1 Remobilisation Des Acquiskeyzoftn.offPas encore d'évaluation
- ProtéomiqueDocument72 pagesProtéomiqueMarwa Sbai100% (1)
- 2020 03 03 CP StressVsImmuniteDocument3 pages2020 03 03 CP StressVsImmuniteNathalieMoyonPas encore d'évaluation
- Download Immunologie Fondamentale Et Immunopathologie 2Nd Edition College Des Enseignants Dimmunologie full chapterDocument61 pagesDownload Immunologie Fondamentale Et Immunopathologie 2Nd Edition College Des Enseignants Dimmunologie full chaptertodd.wimbush786100% (6)
- TD Glucide Medecine 2020 2021 (Série)Document4 pagesTD Glucide Medecine 2020 2021 (Série)Sarah BelbachirPas encore d'évaluation
- IntroductionDocument45 pagesIntroductionYoussef AlaarabiPas encore d'évaluation
- Biotechnologie Agricole Et Transformation de L'Agriculture Ouest-AfricaineDocument29 pagesBiotechnologie Agricole Et Transformation de L'Agriculture Ouest-AfricaineCleaver BrightPas encore d'évaluation
- MicrobiologieDocument30 pagesMicrobiologieangeelfumeroPas encore d'évaluation
- Lesantibiotiquesi 171209223412Document30 pagesLesantibiotiquesi 171209223412minatta100% (1)
- Gautier Jean - L'enfant Ce Glandulaire InconnuDocument206 pagesGautier Jean - L'enfant Ce Glandulaire InconnuNunussePas encore d'évaluation
- Cours15 MYOPATHIES INFLAMMATOIRESDocument51 pagesCours15 MYOPATHIES INFLAMMATOIRESEu VeronicaPas encore d'évaluation
- Chapitre 12 Synthèse Des Protéines (Transcription-Traduction)Document5 pagesChapitre 12 Synthèse Des Protéines (Transcription-Traduction)One Love LifePas encore d'évaluation
- M 23E01 - Cours 01 - CompletDocument36 pagesM 23E01 - Cours 01 - CompletdoliosPas encore d'évaluation
- La CelluleDocument24 pagesLa CelluleMohammed CHENNAOUIPas encore d'évaluation
- L2 Sciences VieDocument2 pagesL2 Sciences VieghfkgkPas encore d'évaluation