Vous aimerez peut-être aussi
- Sommeil: Les Grands Articles d'UniversalisD'EverandSommeil: Les Grands Articles d'UniversalisPas encore d'évaluation
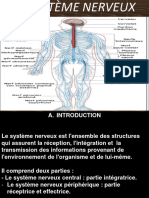
- Resume Sur Le Systeme NerveuxDocument2 pagesResume Sur Le Systeme NerveuxSalam AbdulPas encore d'évaluation
- KULT Feuilles de Personnages - Actions JoueursDocument10 pagesKULT Feuilles de Personnages - Actions JoueursMCRETROGAMING100% (1)
- La Physiologie Du Systeme Nerveux Centrale DR GuedjatiDocument33 pagesLa Physiologie Du Systeme Nerveux Centrale DR GuedjatiMedMan6100% (2)
- Méthodologie de La Recherche Qualitative. Les Questions Clés de La Démarche Compréhensive-2016Document248 pagesMéthodologie de La Recherche Qualitative. Les Questions Clés de La Démarche Compréhensive-2016Achraf MarsoPas encore d'évaluation
- La SomesthésieDocument3 pagesLa SomesthésieZied Bouabene100% (1)
- Physiologie Du Complexe Hypothalamo-HypophysaireDocument5 pagesPhysiologie Du Complexe Hypothalamo-HypophysaireZied Bouabene100% (1)
- Napoli Ieri e Oggi Arr. Lorenzo PuscedduDocument76 pagesNapoli Ieri e Oggi Arr. Lorenzo PuscedduGiuseppe Iuculano100% (2)
- THALAMUSDocument65 pagesTHALAMUSImane BenderazPas encore d'évaluation
- 2-Physiologie Hypothalamo-HypophysaireDocument11 pages2-Physiologie Hypothalamo-HypophysaireNateche NassiraPas encore d'évaluation
- MNE2 Histologie de L Axe Hypotalamo Hypophysaire - + - Epiphyse - + - PancréasDocument10 pagesMNE2 Histologie de L Axe Hypotalamo Hypophysaire - + - Epiphyse - + - PancréasSoheib RahrahPas encore d'évaluation
- Cours Axe Hypothalamo HypophysaireDocument51 pagesCours Axe Hypothalamo HypophysaireSamar OmraniPas encore d'évaluation
- Histologie Spe Üciale UCL Syst EndocrineDocument83 pagesHistologie Spe Üciale UCL Syst EndocrineJosué NAMEGABEPas encore d'évaluation
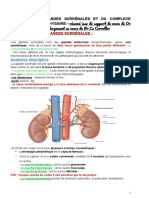
- GLANDES SURRÉNALES22-23-Version LibreDocument7 pagesGLANDES SURRÉNALES22-23-Version LibreLoubna KaouachiPas encore d'évaluation
- Fichier Produit 2195Document25 pagesFichier Produit 2195Raian SabahPas encore d'évaluation
- Neuro Anatomie ??Document170 pagesNeuro Anatomie ??Amirâ ZouânePas encore d'évaluation
- Ronéo N°3Document16 pagesRonéo N°3Djallal HassaniPas encore d'évaluation
- 13.système Nerveux AutonomesDocument18 pages13.système Nerveux AutonomesOumoul KairiPas encore d'évaluation
- 02-Les Hormones Hypothalamo-Hypophysaires (Séance 1+2)Document14 pages02-Les Hormones Hypothalamo-Hypophysaires (Séance 1+2)alimkettouchePas encore d'évaluation
- DiencéphaleDocument6 pagesDiencéphalezainabPas encore d'évaluation
- Eléments de Physiologie - Anatomie Du SNC Et Système Nerveux AutonomeDocument13 pagesEléments de Physiologie - Anatomie Du SNC Et Système Nerveux AutonomeMelPas encore d'évaluation
- Syllabus Neurophysiologie Corrige-1Document131 pagesSyllabus Neurophysiologie Corrige-1Diawara Oumar100% (1)
- Fichier Produit 2272Document9 pagesFichier Produit 2272Amina HasnaouiPas encore d'évaluation
- Chapitre I Organisation Anatomofonctionnelle Du Systeme Nerveux0001Document19 pagesChapitre I Organisation Anatomofonctionnelle Du Systeme Nerveux0001Denisco NkemgnePas encore d'évaluation
- Systeme Nerveux°0Document26 pagesSysteme Nerveux°0salifnchare16Pas encore d'évaluation
- Les Glandes EndocrinesDocument44 pagesLes Glandes EndocrinescepPas encore d'évaluation
- Chapitre IIIDocument5 pagesChapitre IIImhadjyoucef2019Pas encore d'évaluation
- 4) Le Système Nerveux AutonomeDocument4 pages4) Le Système Nerveux AutonomeKassim SankaraPas encore d'évaluation
- Neuroanat-2 0Document83 pagesNeuroanat-2 0seloua kaid100% (1)
- Les Tissus de Protection Du Syste0300me NerveuxDocument6 pagesLes Tissus de Protection Du Syste0300me NerveuxAmel YahiaPas encore d'évaluation
- Chap3 Les Structures Pertinentes Du CerveauDocument3 pagesChap3 Les Structures Pertinentes Du CerveauMahamane Hamissou Salissou Elhadji YahayaPas encore d'évaluation
- Cours Neuroanatomie 2022Document60 pagesCours Neuroanatomie 2022Dr LUCIEN NAWEJPas encore d'évaluation
- Anatomie Du CerveauDocument18 pagesAnatomie Du CerveauSalomon JosephPas encore d'évaluation
- 9 - Physiologie Du Systéme Nerveux AutonomeDocument11 pages9 - Physiologie Du Systéme Nerveux AutonomeAn FelPas encore d'évaluation
- THALAMUS DR GoitaDocument52 pagesTHALAMUS DR GoitaSafia BarraPas encore d'évaluation
- CN1 Neurophysiologie A. Organisation Du Système Nerveux: Comment Peut-On Expliquer Une Telle Évolution?Document23 pagesCN1 Neurophysiologie A. Organisation Du Système Nerveux: Comment Peut-On Expliquer Une Telle Évolution?Oriane plcnPas encore d'évaluation
- Systeme NerveuxDocument82 pagesSysteme Nerveuxmohamedmahfoud198Pas encore d'évaluation
- 01 - Axe HypothalamoDocument10 pages01 - Axe Hypothalamokilo Romeo23Pas encore d'évaluation
- Neuro AnatDocument43 pagesNeuro AnatNouhaila AzirarPas encore d'évaluation
- 1 6 PDFDocument31 pages1 6 PDFcamille.delmotte83Pas encore d'évaluation
- Physiopath Defmot MD 111-130Document18 pagesPhysiopath Defmot MD 111-130Sadi KoukouPas encore d'évaluation
- HYPOPHYSE 2021mesalaDocument48 pagesHYPOPHYSE 2021mesalaMohammed DocPas encore d'évaluation
- Anatomie Et Physiologie Du Système Nerveux-1Document51 pagesAnatomie Et Physiologie Du Système Nerveux-1aurelien choupoPas encore d'évaluation
- Moelle EpiniereDocument6 pagesMoelle Epiniereariane.vasseur29Pas encore d'évaluation
- CM 1 Neurophysiologie - Introduction Et Cellules Gliales N GuéroutDocument82 pagesCM 1 Neurophysiologie - Introduction Et Cellules Gliales N GuéroutfezbejfPas encore d'évaluation
- Cahier TP1 Biomed 2022Document16 pagesCahier TP1 Biomed 2022Tr LwPas encore d'évaluation
- DownloadfileDocument83 pagesDownloadfileAlylia CattPas encore d'évaluation
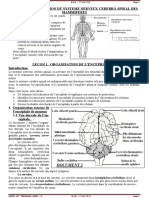
- Theme 1. Organisation Du Systeme Nerveux Cerebro-Spinal Des MammiferesDocument4 pagesTheme 1. Organisation Du Systeme Nerveux Cerebro-Spinal Des MammiferesFatou Diop100% (1)
- PERRIN Neuroanatomie Cours 01 2021Document34 pagesPERRIN Neuroanatomie Cours 01 2021Nassima BoujandirPas encore d'évaluation
- 1 - NeuroanatomieDocument12 pages1 - NeuroanatomieAbdelmoman BoughedadaPas encore d'évaluation
- 6.système NerveuxDocument6 pages6.système NerveuxRahal karimPas encore d'évaluation
- Physiologie Du Systeme NerveuxDocument22 pagesPhysiologie Du Systeme Nerveuxمحمد الأمين ولد عالي ابليلPas encore d'évaluation
- Système NerveuxDocument11 pagesSystème Nerveuxhajar sektiouiPas encore d'évaluation
- 2 2 s1 Systeme NerveuxDocument96 pages2 2 s1 Systeme NerveuxKenza KassabPas encore d'évaluation
- GROUPE N°1, Embryologie Du Système Nerveux Central PDFDocument21 pagesGROUPE N°1, Embryologie Du Système Nerveux Central PDFMondePas encore d'évaluation
- Fascicule SVT TL2 2019-2020 ProfDocument62 pagesFascicule SVT TL2 2019-2020 Profbabacardiop2decPas encore d'évaluation
- Anatomie-Physiologie Du Corps Humain (20-35)Document16 pagesAnatomie-Physiologie Du Corps Humain (20-35)hamria16Pas encore d'évaluation
- Ahh M2decineDocument22 pagesAhh M2decinemalakzerizer2005Pas encore d'évaluation
- AnatomieDocument10 pagesAnatomie1469Pas encore d'évaluation
- Anatmie Externe Du CerveauxDocument8 pagesAnatmie Externe Du CerveauxMedecine DentairePas encore d'évaluation
- Weber - Système Nerveux VégétatifDocument5 pagesWeber - Système Nerveux VégétatifbureljulienPas encore d'évaluation
- MI3 Neurosciences Histologie Du Systeme Nerveux LavabreDocument17 pagesMI3 Neurosciences Histologie Du Systeme Nerveux LavabreDjallal HassaniPas encore d'évaluation
- TD - Système Nerveux - Sans TPDocument11 pagesTD - Système Nerveux - Sans TPFANKELE ALASSANE KONEPas encore d'évaluation
- Lazorthes - Le Système Nerveux PériphériqueDocument400 pagesLazorthes - Le Système Nerveux PériphériqueJoya Gls100% (2)
- De Ribet René-Marcel Les Nerfs CrâniensDocument569 pagesDe Ribet René-Marcel Les Nerfs CrâniensJoya GlsPas encore d'évaluation
- Perlemuter Et Waligora - Cahiers D'anatomie - Abdomen 2Document270 pagesPerlemuter Et Waligora - Cahiers D'anatomie - Abdomen 2Joya Gls33% (3)
- Brizon IV Mucles Du MS - CroquisDocument60 pagesBrizon IV Mucles Du MS - CroquisJoya GlsPas encore d'évaluation
- Brizon VI Vaisseaux Du MS - CroquisDocument26 pagesBrizon VI Vaisseaux Du MS - CroquisJoya GlsPas encore d'évaluation
- Cours D'inittiation À La Recherche 2023Document58 pagesCours D'inittiation À La Recherche 2023Msbbkly DialloPas encore d'évaluation
- Paul FUSTIER - L'interstitiel Et La Fabrique de L'équipeDocument13 pagesPaul FUSTIER - L'interstitiel Et La Fabrique de L'équipeBianca Atodiresei-HüePas encore d'évaluation
- Les Médiations Thérapeutiques Par Lart. Le Réel en JeuDocument326 pagesLes Médiations Thérapeutiques Par Lart. Le Réel en JeuMallis FatiPas encore d'évaluation
- Guide Recrutement Et Accueil Des Administrateurs Obnl - 2022Document21 pagesGuide Recrutement Et Accueil Des Administrateurs Obnl - 2022Martel VictorPas encore d'évaluation
- Mon Carnet - 7eDocument22 pagesMon Carnet - 7edavid.akanPas encore d'évaluation
- Mise en Place de La Séquence 2asDocument1 pageMise en Place de La Séquence 2asKalem RaniaPas encore d'évaluation
- Présentation Cours GoffmanDocument63 pagesPrésentation Cours GoffmanAmina MedPas encore d'évaluation
- L Influence de Kant Sur La Conception Arendtienne de La Libert PolitiqueDocument2 pagesL Influence de Kant Sur La Conception Arendtienne de La Libert PolitiqueMounirPas encore d'évaluation
- Cours - La Communication Non ViolenteDocument2 pagesCours - La Communication Non ViolenteBriki MaramPas encore d'évaluation
- Feuille de Paie 4Document2 pagesFeuille de Paie 4coolbil123lovePas encore d'évaluation
- Triple JeDocument157 pagesTriple Jepeut etrePas encore d'évaluation
- Communication en EntrepriseDocument14 pagesCommunication en EntrepriseyasminePas encore d'évaluation
- Adoramus Te, Christe - DuboisDocument1 pageAdoramus Te, Christe - DuboisVox CantorisPas encore d'évaluation
- STRATEGIES Les Facilitateurs Pédagogiques.Document8 pagesSTRATEGIES Les Facilitateurs Pédagogiques.Daniel MoukouriPas encore d'évaluation