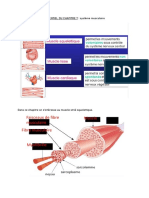
Vous aimerez peut-être aussi
- Muscles: Les Grands Articles d'UniversalisD'EverandMuscles: Les Grands Articles d'UniversalisPas encore d'évaluation
- Tissu Musculaire PDFDocument58 pagesTissu Musculaire PDFayoubPas encore d'évaluation
- Physiologie Musculaire 2020Document47 pagesPhysiologie Musculaire 2020drahlemtrifPas encore d'évaluation
- 2 S7 Cours 2Document44 pages2 S7 Cours 2Chabane OubarechePas encore d'évaluation
- Physiologie Animale Cours 15Document168 pagesPhysiologie Animale Cours 15g69wwpfw5kPas encore d'évaluation
- Physio1an 05 Physiologie Du Muscle Strie-KermicheDocument25 pagesPhysio1an 05 Physiologie Du Muscle Strie-KermicheSamwil SUANONPas encore d'évaluation
- Contracté - Relâché, Encore Une Fois. - François RosenzweigDocument102 pagesContracté - Relâché, Encore Une Fois. - François RosenzweigOlivia ManceauPas encore d'évaluation
- Tissu Musculaire MeksemDocument45 pagesTissu Musculaire MeksemNo Comment.Pas encore d'évaluation
- BP 2 Boucherie - Le Muscle-CorrectionDocument13 pagesBP 2 Boucherie - Le Muscle-CorrectionThierry Pano Doubleface DucosPas encore d'évaluation
- Syst MusculaireDocument74 pagesSyst Musculaireberthelotsacha85Pas encore d'évaluation
- 03 - 2 - Proprietes Mecaniques Du Muscle - ContractionDocument30 pages03 - 2 - Proprietes Mecaniques Du Muscle - ContractionGloria KitonPas encore d'évaluation
- CM - Composition Et Fonction D'un MuscleDocument3 pagesCM - Composition Et Fonction D'un MuscleBassetPas encore d'évaluation
- Système MusculaireDocument16 pagesSystème MusculaireMustapha BriachePas encore d'évaluation
- Le Muscle NormalDocument41 pagesLe Muscle NormalHelena KenguePas encore d'évaluation
- 03 - 1 - Proprietes Mecaniques Du Muscle - GeneralitesDocument32 pages03 - 1 - Proprietes Mecaniques Du Muscle - GeneralitesGloria KitonPas encore d'évaluation
- Le Système Musculaire Et Le DopageDocument34 pagesLe Système Musculaire Et Le DopageArnaud DimigouPas encore d'évaluation
- Projet EMG (ELECROMYOGRAPHE) FinaliséDocument64 pagesProjet EMG (ELECROMYOGRAPHE) FinaliséAssia Saoudi100% (3)
- tp7 CorrigeDocument4 pagestp7 CorrigeEmmanuel KoffiPas encore d'évaluation
- MuscleDocument15 pagesMuscleMaximePas encore d'évaluation
- Le Systeme Musculaire Serie D Exercices Word 1Document11 pagesLe Systeme Musculaire Serie D Exercices Word 1zinebsemlali2009Pas encore d'évaluation
- Cours Myologie PsychomotDocument7 pagesCours Myologie PsychomotRazenAlgPas encore d'évaluation
- Poly Physiologie Cellulaire - Contraction Musculaire 2006-2007Document91 pagesPoly Physiologie Cellulaire - Contraction Musculaire 2006-2007roger kinaPas encore d'évaluation
- ESSENTIEL Moitié Des ChapitresDocument18 pagesESSENTIEL Moitié Des ChapitresLucie ImbertPas encore d'évaluation
- Cours 5 Tissu Musculaire PDFDocument5 pagesCours 5 Tissu Musculaire PDFOmar KodbePas encore d'évaluation
- Le Systeme Musculaire Resume de Cours MaDocument1 pageLe Systeme Musculaire Resume de Cours MaAymane JiraouiPas encore d'évaluation
- Le Muscle Strie SquelettiqueDocument3 pagesLe Muscle Strie SquelettiqueBathie SarrPas encore d'évaluation
- Les Tissus MusculairesDocument56 pagesLes Tissus MusculairesIdrissou Fmsb100% (2)
- Devoir PhysiologieDocument18 pagesDevoir PhysiologieBetschbel DorcéPas encore d'évaluation
- Système Musculaire 2020Document22 pagesSystème Musculaire 2020Sarah OuajnatiPas encore d'évaluation
- Physiologie de L'appareil Locomoteur-Le Système MusculaireDocument6 pagesPhysiologie de L'appareil Locomoteur-Le Système MusculaireEmy DesreumauxPas encore d'évaluation
- MOTRICITE Par 2 DiapDocument79 pagesMOTRICITE Par 2 DiapRaphaël BikoahPas encore d'évaluation
- Tissue Musculaire - para Clase 2Document39 pagesTissue Musculaire - para Clase 2tntanime00Pas encore d'évaluation
- 1.physiologie Musculaire 2023Document59 pages1.physiologie Musculaire 2023aminezebar3Pas encore d'évaluation
- ch10 Solutionnaire 1569538426Document12 pagesch10 Solutionnaire 1569538426Wilfried KouaoPas encore d'évaluation
- Le Fonctionnement Du Muscle Squelettique: Magazine de Sciences de La Vie Et de La TerreDocument28 pagesLe Fonctionnement Du Muscle Squelettique: Magazine de Sciences de La Vie Et de La TerreSifddin GuidPas encore d'évaluation
- Système Musculaire 2021Document24 pagesSystème Musculaire 2021Yassine YacinePas encore d'évaluation
- Cours (Système Musculaire)Document3 pagesCours (Système Musculaire)usernoone2001Pas encore d'évaluation
- Tissu MusculaireDocument14 pagesTissu MusculaireSana JebbarPas encore d'évaluation
- Système Musculaire 3AC PR - naciRI IssameDocument6 pagesSystème Musculaire 3AC PR - naciRI IssameElhoussainePas encore d'évaluation
- Système Musculaire 3AC PR - NACIRI Issame (WWW - Pc1.ma)Document6 pagesSystème Musculaire 3AC PR - NACIRI Issame (WWW - Pc1.ma)Mary JohanaPas encore d'évaluation
- 5 La MyologieDocument19 pages5 La MyologieYou NesPas encore d'évaluation
- Le Système MusculaireDocument9 pagesLe Système MusculaireTahiri AbdelilahPas encore d'évaluation
- Système MusculaireDocument7 pagesSystème MusculaireAmine WlPas encore d'évaluation
- Boutonnat Jean p01 PDFDocument61 pagesBoutonnat Jean p01 PDFMoncef PechaPas encore d'évaluation
- 7 Tissu Musculaire Polycope 2020Document15 pages7 Tissu Musculaire Polycope 2020randomPas encore d'évaluation
- CM Logique Musculo-SquelettiqueDocument50 pagesCM Logique Musculo-Squelettiquemarine falzeiPas encore d'évaluation
- La MyologieDocument6 pagesLa MyologieBilly Bel Vocale MesidorPas encore d'évaluation
- Les MusculesDocument5 pagesLes Musculesnouhailabenallal768Pas encore d'évaluation
- Cours MuscleDocument64 pagesCours Musclezerkarim222Pas encore d'évaluation
- Cours Du Tissu Musculaire 2023Document9 pagesCours Du Tissu Musculaire 2023agag.salahPas encore d'évaluation
- Sequence 2 Cours Svteehb Tle D 2023Document49 pagesSequence 2 Cours Svteehb Tle D 2023Brice ONKELPas encore d'évaluation
- 7 - Les Tissus MusculairesDocument6 pages7 - Les Tissus MusculairesHayfa Ben taherPas encore d'évaluation
- Le Muscle: SquelettiqueDocument8 pagesLe Muscle: SquelettiqueHamza LiberadosPas encore d'évaluation
- Chapitre 4 Et Chapitre 5 Muscles Lisse Et Cardiaque.Document14 pagesChapitre 4 Et Chapitre 5 Muscles Lisse Et Cardiaque.habessongPas encore d'évaluation
- Les MusclesDocument4 pagesLes Muscleschiha hocinePas encore d'évaluation
- TSpe T3B1 CoursDocument5 pagesTSpe T3B1 CoursFranckyscocom Gokpan Enfant BeniPas encore d'évaluation
- P Motricité (C. Quairiaux) - 26.03.21Document5 pagesP Motricité (C. Quairiaux) - 26.03.21nasten16fhPas encore d'évaluation
- Contraction Musculaire CouleurDocument286 pagesContraction Musculaire Couleurmatis47215100% (1)
- 3 Myologie DEF LightDocument109 pages3 Myologie DEF LightCherbiti Mohammed AminePas encore d'évaluation
- 4) - Le Système MusculaireDocument18 pages4) - Le Système Musculairemaria bounouaPas encore d'évaluation
- Reunion ETR 2009-09-19Document18 pagesReunion ETR 2009-09-19patricePas encore d'évaluation
- Xerox Scan - 2506201351Document4 pagesXerox Scan - 2506201351Fafa FafaPas encore d'évaluation
- Semio Hemato19-HemogrammeDocument6 pagesSemio Hemato19-HemogrammebelkassemPas encore d'évaluation
- Unite 2 Les TissusDocument41 pagesUnite 2 Les TissusFaycal BennaniPas encore d'évaluation
- Physiologie Et Exploration de LDocument8 pagesPhysiologie Et Exploration de LAymenPas encore d'évaluation
- Cours Hemato Cpfim&Document5 pagesCours Hemato Cpfim&Nicodeme SodeaPas encore d'évaluation
- مركز هبةDocument65 pagesمركز هبةprint housePas encore d'évaluation
- LNME Cameroun 2017 ReactifsDocument7 pagesLNME Cameroun 2017 ReactifsDjoufackPas encore d'évaluation
- Copie de 3.3 TissusDocument25 pagesCopie de 3.3 TissusAmber Stolz élèvePas encore d'évaluation
- COURS IH LAM3L Séquence 1 Ag ABO 04 03 18 Séquence 1Document20 pagesCOURS IH LAM3L Séquence 1 Ag ABO 04 03 18 Séquence 1rostandtchangouePas encore d'évaluation
- 98 549Document5 pages98 549Tewfik TGrPas encore d'évaluation
- PancytopéniesDocument8 pagesPancytopénieserwin NZOLPas encore d'évaluation
- TD HG DevoirsDocument4 pagesTD HG Devoirsyennefer vengerbergPas encore d'évaluation
- Drepanocytose - CopieDocument69 pagesDrepanocytose - Copiesadibarry01Pas encore d'évaluation
- TP6 Tissus MusculairesDocument7 pagesTP6 Tissus MusculairesNihal KrikaPas encore d'évaluation
- 03 - 1 - Proprietes Mecaniques Du Muscle - GeneralitesDocument32 pages03 - 1 - Proprietes Mecaniques Du Muscle - GeneralitesGloria KitonPas encore d'évaluation
- Physiologie de La Pression IntracranienneDocument32 pagesPhysiologie de La Pression IntracranienneRyma TriekPas encore d'évaluation
- Les Epitheliums 2021.Document89 pagesLes Epitheliums 2021.Hanne ZamelPas encore d'évaluation
- Frequence Des Anemies Au Milieu HospitailierDocument50 pagesFrequence Des Anemies Au Milieu HospitailierDjaouida DjiralPas encore d'évaluation
- Etats de Choc Hemorragiques 2020Document58 pagesEtats de Choc Hemorragiques 2020abdo217100% (1)
- Manuel de Procedures RNL SNDocument97 pagesManuel de Procedures RNL SNstephane legrandPas encore d'évaluation
- Groupe Sanguin CompatibiliteDocument2 pagesGroupe Sanguin CompatibiliteSovac2019 SovacPas encore d'évaluation
- Questions-Réponses CrâneDocument8 pagesQuestions-Réponses CrâneAly GuindoPas encore d'évaluation
- CR Résultats D'examens BiologiquesDocument3 pagesCR Résultats D'examens BiologiquesMor Talla DiouckPas encore d'évaluation
- 2001 A 2 Lipome LiposarcomeDocument12 pages2001 A 2 Lipome LiposarcomeVladlena CaraimanPas encore d'évaluation
- 18 Hs II Dr. S. MetatlaDocument50 pages18 Hs II Dr. S. MetatlaDr BENOUADFELPas encore d'évaluation
- 17 ch08 085 100 9782294769580Document19 pages17 ch08 085 100 9782294769580ionefoiznPas encore d'évaluation
- TD 3 CorrigéDocument10 pagesTD 3 CorrigéIdrissou Fmsb0% (1)
- 5 Groupes Sanguins, Transfusion Sanguine Et Accidents de La TransfusionDocument5 pages5 Groupes Sanguins, Transfusion Sanguine Et Accidents de La TransfusionkhiralouibiPas encore d'évaluation
- Cours DhemobiologieDocument2 pagesCours DhemobiologiedorcasmerveillegfotsoPas encore d'évaluation
- La vie des abeilles: Prix Nobel de littératureD'EverandLa vie des abeilles: Prix Nobel de littératureÉvaluation : 4 sur 5 étoiles4/5 (41)
- Histoire de la psychologie scientifique: De la naissance de la psychologie à la neuropsychologie et aux champs d'application les plus actuelsD'EverandHistoire de la psychologie scientifique: De la naissance de la psychologie à la neuropsychologie et aux champs d'application les plus actuelsPas encore d'évaluation
- Hypnose & affirmations positives pour la richesse, l’abondance et la réussite (21 jours pour un nouveau vous)D'EverandHypnose & affirmations positives pour la richesse, l’abondance et la réussite (21 jours pour un nouveau vous)Évaluation : 5 sur 5 étoiles5/5 (1)
- Cahier de jeux de stimulation cognitive: Sujets Alzheimer, désorientés, démences, amnésiesD'EverandCahier de jeux de stimulation cognitive: Sujets Alzheimer, désorientés, démences, amnésiesPas encore d'évaluation
- Introduction à la psychologie des émotions: De Darwin aux neurosciences: découvrir les émotions et leur mode de fonctionnementD'EverandIntroduction à la psychologie des émotions: De Darwin aux neurosciences: découvrir les émotions et leur mode de fonctionnementPas encore d'évaluation
- Neuropsychologie: Les bases théoriques et pratiques du domaine d'étude (psychologie pour tous)D'EverandNeuropsychologie: Les bases théoriques et pratiques du domaine d'étude (psychologie pour tous)Pas encore d'évaluation
- Améliorer votre mémoire: Un Guide pour l'augmentation de la puissance du cerveau, utilisant des techniques et méthodesD'EverandAméliorer votre mémoire: Un Guide pour l'augmentation de la puissance du cerveau, utilisant des techniques et méthodesÉvaluation : 5 sur 5 étoiles5/5 (2)
- Harmonisation Energétique des Lieux: Habitat et haut-lieux sacrés 2020D'EverandHarmonisation Energétique des Lieux: Habitat et haut-lieux sacrés 2020Évaluation : 2.5 sur 5 étoiles2.5/5 (3)
- Le TDAH chez l’adulte : Comment reconnaître & faire face au TDAH chez l’adulte en 30 étapes faciles.D'EverandLe TDAH chez l’adulte : Comment reconnaître & faire face au TDAH chez l’adulte en 30 étapes faciles.Pas encore d'évaluation
- Géologie de l'Amérique: Les Grands Articles d'UniversalisD'EverandGéologie de l'Amérique: Les Grands Articles d'UniversalisPas encore d'évaluation
- Hypersensible Et alors ?: Un tempérament expliquéD'EverandHypersensible Et alors ?: Un tempérament expliquéÉvaluation : 5 sur 5 étoiles5/5 (2)
- La thérapie libre: La Thérapie de LIBération et Résilience EmotionnelleD'EverandLa thérapie libre: La Thérapie de LIBération et Résilience EmotionnelleÉvaluation : 5 sur 5 étoiles5/5 (1)
- La psychologie de l'anxiété: La connaître pour comprendre ses mécanismes de fonctionnementD'EverandLa psychologie de l'anxiété: La connaître pour comprendre ses mécanismes de fonctionnementPas encore d'évaluation
- Pourquoi suis-je différent?: Accepter et découvrir la puissance de l'hypersensibilitéD'EverandPourquoi suis-je différent?: Accepter et découvrir la puissance de l'hypersensibilitéPas encore d'évaluation
- Autisme - La boîte à outils: Stratégies et techniques pour accompagner un enfant autisteD'EverandAutisme - La boîte à outils: Stratégies et techniques pour accompagner un enfant autistePas encore d'évaluation
- Le CODE DE DIEU: Le secret de notre passé, la promesse de notre avenirD'EverandLe CODE DE DIEU: Le secret de notre passé, la promesse de notre avenirÉvaluation : 4 sur 5 étoiles4/5 (23)
- Mes tempêtes intérieures: 25 témoignages porteurs d'espoir sur la santé mentaleD'EverandMes tempêtes intérieures: 25 témoignages porteurs d'espoir sur la santé mentalePas encore d'évaluation
- Le Pouvoir De L'Autodiscipline: L’art de tout accomplir sans limiteD'EverandLe Pouvoir De L'Autodiscipline: L’art de tout accomplir sans limiteÉvaluation : 4 sur 5 étoiles4/5 (16)
- Qu'arrive t'il a la terre arable ?: Le problème de l'érosion du sol.D'EverandQu'arrive t'il a la terre arable ?: Le problème de l'érosion du sol.Évaluation : 5 sur 5 étoiles5/5 (1)
- La motivation: Voyage au cœur des comportements motivés, de l'étude des processus internes aux théories neuropsychologiques les plus récentesD'EverandLa motivation: Voyage au cœur des comportements motivés, de l'étude des processus internes aux théories neuropsychologiques les plus récentesPas encore d'évaluation
- Connaître ses réactions émotionnelles, pour maîtriser ses émotionsD'EverandConnaître ses réactions émotionnelles, pour maîtriser ses émotionsPas encore d'évaluation
- Atlas permanent de l'Union européenne: 5e édition revue et augmentéeD'EverandAtlas permanent de l'Union européenne: 5e édition revue et augmentéePas encore d'évaluation
- Transition énergétique ET Changement climatique: Développements et Perspectives d'AvenirD'EverandTransition énergétique ET Changement climatique: Développements et Perspectives d'AvenirPas encore d'évaluation