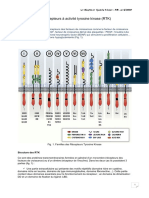
Vous aimerez peut-être aussi
- Inhibition de La Voie de MEKDocument11 pagesInhibition de La Voie de MEKtaoufik akabliPas encore d'évaluation
- INTRODUCTIONDocument7 pagesINTRODUCTIONmy computerPas encore d'évaluation
- UntithggDocument18 pagesUntithggLoubna MichaPas encore d'évaluation
- UntitledtdugogrDocument3 pagesUntitledtdugogrLoubna MichaPas encore d'évaluation
- Article02 21 22Document9 pagesArticle02 21 22Moncef CherifPas encore d'évaluation
- Resistance To Thyroid Hormone: SFE Paris 2013 / Annales D'endocrinologie 74 (2013) 238-239Document1 pageResistance To Thyroid Hormone: SFE Paris 2013 / Annales D'endocrinologie 74 (2013) 238-239henry hernandezPas encore d'évaluation
- Cancero Dig 2010 1 038 042-With-Cover-Page-V2Document6 pagesCancero Dig 2010 1 038 042-With-Cover-Page-V2Loubna MichaPas encore d'évaluation
- TD Signal 2023Document10 pagesTD Signal 2023salym.mourid84Pas encore d'évaluation
- Ion Superoxyde Et PhagocytesDocument2 pagesIon Superoxyde Et PhagocytesJean-Loïc BauchetPas encore d'évaluation
- MS 1994 6-7Document8 pagesMS 1994 6-7Loubna MichaPas encore d'évaluation
- Cours RéglationDocument33 pagesCours RéglationBADzPas encore d'évaluation
- CortisolDocument3 pagesCortisolAcide OléiquePas encore d'évaluation
- Récepteurs À Activité Tyrosine Kinase DefDocument12 pagesRécepteurs À Activité Tyrosine Kinase DefFatima ZahraPas encore d'évaluation
- Travail BioinformatiqueDocument13 pagesTravail Bioinformatiqueriahi aymenPas encore d'évaluation
- Article A Analyser FRDocument20 pagesArticle A Analyser FRasmabac21Pas encore d'évaluation
- 01 C Interaction Ligand RécepteurDocument39 pages01 C Interaction Ligand RécepteurKhelifi SarraPas encore d'évaluation
- 2.-Biosynthèse Des ProtéinesDocument65 pages2.-Biosynthèse Des Protéinesmido benPas encore d'évaluation
- GTSE1 Est Impliqué Dans La Progression Du Cancer Du Sein de Manière Dépendante de La Mutation p53Document45 pagesGTSE1 Est Impliqué Dans La Progression Du Cancer Du Sein de Manière Dépendante de La Mutation p53Haby KonatéPas encore d'évaluation
- GB3!09!10-Signalisation-Cours E. Macia Généralités Et RCPGDocument118 pagesGB3!09!10-Signalisation-Cours E. Macia Généralités Et RCPGlobnaPas encore d'évaluation
- 3-Régulation de L - Expression Des Gènes N.MDocument3 pages3-Régulation de L - Expression Des Gènes N.Msevda mouviePas encore d'évaluation
- Fin OkDocument7 pagesFin Okposteraq374Pas encore d'évaluation
- Qe BiocellDocument9 pagesQe BiocellASSIA EL MOUSTAID100% (1)
- Résumé Chapitre 4 Biochimie Cellulaire Et FonctionnelleDocument30 pagesRésumé Chapitre 4 Biochimie Cellulaire Et Fonctionnelledraboissouf174Pas encore d'évaluation
- TD #5: Synthèse Des Protéines de Sécrétion Au Niveau Du REGDocument7 pagesTD #5: Synthèse Des Protéines de Sécrétion Au Niveau Du REGAy ManPas encore d'évaluation
- Génétique Et ÉpigénétiqueDocument41 pagesGénétique Et ÉpigénétiqueAdli OthmanePas encore d'évaluation
- Medsci 2012285 P 465Document4 pagesMedsci 2012285 P 465KougangPas encore d'évaluation
- Biologie Moléculaire Chapitre 1 T Procaryote M1Document8 pagesBiologie Moléculaire Chapitre 1 T Procaryote M1اميرة في عرش ابيPas encore d'évaluation
- Ronéo Ronéos Biologie Biologie 4 Fini 2022Document12 pagesRonéo Ronéos Biologie Biologie 4 Fini 2022lialazzaraPas encore d'évaluation
- Biologie Moléculaire TD 03Document29 pagesBiologie Moléculaire TD 03biotech micro100% (1)
- MED1 TransductionDocument105 pagesMED1 TransductionSamir GhoualiPas encore d'évaluation
- III. Mécanismes Moléculaires de L'expression de L'information GénétiqueDocument54 pagesIII. Mécanismes Moléculaires de L'expression de L'information GénétiqueNaima BouhmaidaPas encore d'évaluation
- ChapIV Signalisation Et RégulationDocument37 pagesChapIV Signalisation Et RégulationMamadou lamine DiattaPas encore d'évaluation
- Les Récepteurs À Activité Tyrosine KinaseDocument10 pagesLes Récepteurs À Activité Tyrosine KinaseFatima Zahra100% (1)
- Cancers 1500Document29 pagesCancers 1500Mounirou BaparapePas encore d'évaluation
- Biologie Moléculaire Chapitre 3 REGULATION M1 1Document8 pagesBiologie Moléculaire Chapitre 3 REGULATION M1 1اميرة في عرش ابيPas encore d'évaluation
- Exposé Du Groupe 9Document35 pagesExposé Du Groupe 9Grâce KABOREPas encore d'évaluation
- 2 OukalDocument49 pages2 OukalDidier chanelPas encore d'évaluation
- Recepteur de L'insuline G1Document12 pagesRecepteur de L'insuline G1페 리알Pas encore d'évaluation
- PFE SemiDocument37 pagesPFE SemiJeremy OueretPas encore d'évaluation
- Cours BMDocument22 pagesCours BMNidhaloo NidPas encore d'évaluation
- Thérapies Ciblées 231010 165853Document5 pagesThérapies Ciblées 231010 165853moussaoued493Pas encore d'évaluation
- Dysfonctionnement Du Cycle Cellulaire Et PathologiesDocument20 pagesDysfonctionnement Du Cycle Cellulaire Et Pathologiesmina ycPas encore d'évaluation
- Cellules Souches PluripotentesDocument7 pagesCellules Souches Pluripotentesmariem.bourouniaPas encore d'évaluation
- Les Thérapeutiques Ciblées Du Cancer de Vessie Localement Avancé Et/ou MétastatiqueDocument11 pagesLes Thérapeutiques Ciblées Du Cancer de Vessie Localement Avancé Et/ou MétastatiqueBoutaina AddoumPas encore d'évaluation
- TRANSCRIPTION S4 ÉtudiantsDocument6 pagesTRANSCRIPTION S4 ÉtudiantsNiavo HeriniriantsoaPas encore d'évaluation
- Isoprenyl Carboxyl Methyltransferase Inhibitors - A Brief Review Including Recent Patents - PMCDocument24 pagesIsoprenyl Carboxyl Methyltransferase Inhibitors - A Brief Review Including Recent Patents - PMCMouhcine MohammedPas encore d'évaluation
- Transcription 21 - 11 - 2023Document55 pagesTranscription 21 - 11 - 2023Elmahdi BOUHFIDPas encore d'évaluation
- Abc-304319-Accumulation de La Proteine p53 Et Surexpression Du Recepteur Her3 Dans Le Cancer Colorectal - W8uYVn8AAQEAAGL4opUAAAAC-aDocument7 pagesAbc-304319-Accumulation de La Proteine p53 Et Surexpression Du Recepteur Her3 Dans Le Cancer Colorectal - W8uYVn8AAQEAAGL4opUAAAAC-aWassCANPas encore d'évaluation
- Analyse Partie Disscussion (Finale)Document5 pagesAnalyse Partie Disscussion (Finale)kaouthar boumaichePas encore d'évaluation
- Recepteur A Activite Serine Threonine KinaseDocument11 pagesRecepteur A Activite Serine Threonine KinaseFatima ZahraPas encore d'évaluation
- Genetika Maligne CelijeDocument34 pagesGenetika Maligne CelijejovanaPas encore d'évaluation
- Mecanismes Moleculaires Controlant Le CyDocument21 pagesMecanismes Moleculaires Controlant Le CymohamedPas encore d'évaluation
- Chap 7Document22 pagesChap 7Ayoub LilouPas encore d'évaluation
- Compartiments Intracell Et Tri Des ProtéinesDocument43 pagesCompartiments Intracell Et Tri Des Protéinesa29921Pas encore d'évaluation
- SignalisationDocument35 pagesSignalisationpathcell bioPas encore d'évaluation
- Thérapies CibléesDocument10 pagesThérapies Cibléesceliaboizeau09Pas encore d'évaluation
- Med Genetique-Transcription EucaryotesDocument20 pagesMed Genetique-Transcription EucaryotesRacha MalakPas encore d'évaluation
- Ziani Module Biologie Moléculaire Et Génie Génétique L3 MicobiologieDocument122 pagesZiani Module Biologie Moléculaire Et Génie Génétique L3 MicobiologieOwassiPas encore d'évaluation
- Efusi Pleura MalignaDocument10 pagesEfusi Pleura MalignaEVa ApianiPas encore d'évaluation
- Communications Et Signalisations Cellulaires 4 Ed - SommaireDocument28 pagesCommunications Et Signalisations Cellulaires 4 Ed - SommaireAyoub Lilou100% (1)
- Les Perturbateurs Endocriniens Quel Risque Pour La SantéDocument10 pagesLes Perturbateurs Endocriniens Quel Risque Pour La SantéjamjamalPas encore d'évaluation
- Signalisation CellulaireDocument47 pagesSignalisation CellulaireJamaa Bouzkri0% (1)
- BiocellulaireDocument4 pagesBiocellulairecarlio12Pas encore d'évaluation
- Biologie Végétale: Croissance Et DéveloppementDocument24 pagesBiologie Végétale: Croissance Et DéveloppementGOUNOUPas encore d'évaluation
- L Essentiel de La Biologie Cellulaire 3 Ed Chapitre16Document18 pagesL Essentiel de La Biologie Cellulaire 3 Ed Chapitre16Vayo SonyPas encore d'évaluation
- CytokinDocument18 pagesCytokinwassim hamdikenePas encore d'évaluation
- Les Récepteurs À Activité Tyrosine KinaseDocument10 pagesLes Récepteurs À Activité Tyrosine KinaseFatima Zahra100% (1)
- TravailDocument2 pagesTravailBerthe Aristide Ngue NkongoPas encore d'évaluation
- Cours 1 Généralités Sur La Signalisation Cellulaire 1Document18 pagesCours 1 Généralités Sur La Signalisation Cellulaire 1abdou.djat75Pas encore d'évaluation
- Biochimie Des Hormones Et Leurs Mécanismes D'action-Récepteurs Membranaires-1Document22 pagesBiochimie Des Hormones Et Leurs Mécanismes D'action-Récepteurs Membranaires-1AGUEZLANE NOURPas encore d'évaluation
- Physiopathologie de L'insulinorésistance 2019Document10 pagesPhysiopathologie de L'insulinorésistance 2019Lilo LiliPas encore d'évaluation
- Interaction Ligand RécepteurDocument6 pagesInteraction Ligand RécepteurFaculté De Médecine BécharPas encore d'évaluation
- le Signal de Transduction MembranaireDocument25 pagesle Signal de Transduction MembranairekerbabdjamilaPas encore d'évaluation
- Défaut de Migration Des NeutrophilesDocument14 pagesDéfaut de Migration Des NeutrophilesDjamaleddine FahPas encore d'évaluation
- Abc-304319-Accumulation de La Proteine p53 Et Surexpression Du Recepteur Her3 Dans Le Cancer Colorectal - W8uYVn8AAQEAAGL4opUAAAAC-aDocument7 pagesAbc-304319-Accumulation de La Proteine p53 Et Surexpression Du Recepteur Her3 Dans Le Cancer Colorectal - W8uYVn8AAQEAAGL4opUAAAAC-aWassCANPas encore d'évaluation
- S1Cours de Pharmacologie Générale M1 Physio BoucennaDocument11 pagesS1Cours de Pharmacologie Générale M1 Physio BoucennaDaniel NdPas encore d'évaluation
- Les CytokinesDocument47 pagesLes CytokinesOusmane SyllaPas encore d'évaluation
- LivreCancer PDFDocument136 pagesLivreCancer PDFAdèle N.Pas encore d'évaluation
- Molécules D'adhérence Cours L1Document3 pagesMolécules D'adhérence Cours L1safemindPas encore d'évaluation
- Embryologie Humaine: de LarsenDocument28 pagesEmbryologie Humaine: de LarsenYu-oh Crysis100% (1)
- These C.jousseDocument174 pagesThese C.jousseMina BekPas encore d'évaluation
- Cour4 Expression D'antigènes, Marqueurs de Virulence Et de Récepteurs CellulairesDocument6 pagesCour4 Expression D'antigènes, Marqueurs de Virulence Et de Récepteurs CellulairesJoie De VivrePas encore d'évaluation
- THPH 2016 MUZIN QuentinDocument287 pagesTHPH 2016 MUZIN QuentinChristian BriardPas encore d'évaluation
- Corps Humain 2H UE 2.2 2017Document43 pagesCorps Humain 2H UE 2.2 2017Nambinintsoa HeriniainaPas encore d'évaluation
- Cours Biologie ApoptoseDocument11 pagesCours Biologie Apoptoseoussama bousseloubPas encore d'évaluation
- 1 Introduction A La Physiologie Des Glandes EndocrinesDocument16 pages1 Introduction A La Physiologie Des Glandes EndocrinesAlina Topor Psi-MedicalPas encore d'évaluation
- Genetique Moleculaire Humaine 4 Ed - SommaireDocument26 pagesGenetique Moleculaire Humaine 4 Ed - SommaireDJIL BENPas encore d'évaluation