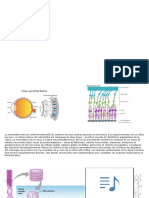
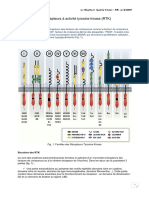
Vous aimerez peut-être aussi
- 11 - Chap11-Les Différents Modes D'action Des HormonesDocument6 pages11 - Chap11-Les Différents Modes D'action Des HormonesYassine HassanePas encore d'évaluation
- Définition de La Protéine GDocument4 pagesDéfinition de La Protéine GAsma MhPas encore d'évaluation
- UntitledtdugogrDocument3 pagesUntitledtdugogrLoubna MichaPas encore d'évaluation
- Interaction Ligand RecepteurDocument3 pagesInteraction Ligand RecepteurAmina MnPas encore d'évaluation
- MED1 TransductionDocument105 pagesMED1 TransductionSamir GhoualiPas encore d'évaluation
- Interactions Ligand - Recepteur 2023-2024Document29 pagesInteractions Ligand - Recepteur 2023-2024khaldiniheddoc23Pas encore d'évaluation
- CortisolDocument3 pagesCortisolAcide OléiquePas encore d'évaluation
- Les Récepteurs À Activité Tyrosine KinaseDocument10 pagesLes Récepteurs À Activité Tyrosine KinaseFatima Zahra100% (1)
- Communication CellulaireDocument22 pagesCommunication CellulaireAmïn Fïlalï100% (2)
- 01 C Interaction Ligand RécepteurDocument39 pages01 C Interaction Ligand RécepteurKhelifi SarraPas encore d'évaluation
- Recepteur de L'insuline G1Document12 pagesRecepteur de L'insuline G1페 리알Pas encore d'évaluation
- Neurophysiologie Générale - Les Transmissions Synaptiques Lentes Et Les Cellules GlialesDocument10 pagesNeurophysiologie Générale - Les Transmissions Synaptiques Lentes Et Les Cellules GlialesanaischapellzPas encore d'évaluation
- Physio1an-Interactions Ligand Recepteur2021bougrida BoussoufDocument24 pagesPhysio1an-Interactions Ligand Recepteur2021bougrida BoussoufGI 2 Session 2018-2019Pas encore d'évaluation
- G-Protein in Cell Signaling - IntroductionDocument11 pagesG-Protein in Cell Signaling - Introductiontaoufik akabliPas encore d'évaluation
- La Cellule AnimaleDocument10 pagesLa Cellule AnimaleAmanda LaihongtingPas encore d'évaluation
- Récepteurs Couplés Au Protéines G Partie 2Document7 pagesRécepteurs Couplés Au Protéines G Partie 2Fatima ZahraPas encore d'évaluation
- Récepteurs À Activité Tyrosine Kinase DefDocument12 pagesRécepteurs À Activité Tyrosine Kinase DefFatima ZahraPas encore d'évaluation
- UE2-Intégration - 13 - Communication-IntercellulaireDocument17 pagesUE2-Intégration - 13 - Communication-Intercellulairesun-nee-chan9Pas encore d'évaluation
- GB3!09!10-Signalisation-Cours E. Macia Généralités Et RCPGDocument118 pagesGB3!09!10-Signalisation-Cours E. Macia Généralités Et RCPGlobnaPas encore d'évaluation
- Interaction Ligand RécepteurDocument6 pagesInteraction Ligand RécepteurFaculté De Médecine BécharPas encore d'évaluation
- Recepteurs Associes A Une Tyrosine KinaseDocument13 pagesRecepteurs Associes A Une Tyrosine KinaseFatima ZahraPas encore d'évaluation
- 01 - La Communication IntercellulaireDocument8 pages01 - La Communication IntercellulaireHadil BenslamaPas encore d'évaluation
- Cours 8 A GLYCOLYSEDocument15 pagesCours 8 A GLYCOLYSELina BernoussiPas encore d'évaluation
- Cibles Moléculaires Et Élaboration de La Réponse 2024Document3 pagesCibles Moléculaires Et Élaboration de La Réponse 2024kaouaneamdjadPas encore d'évaluation
- 2023-3BB - Classes RCDocument36 pages2023-3BB - Classes RCIsabella AlmeidaPas encore d'évaluation
- INTRODUCTIONDocument7 pagesINTRODUCTIONmy computerPas encore d'évaluation
- Bioch Pharm2an-Metabolisme GlucidesDocument28 pagesBioch Pharm2an-Metabolisme GlucidesmaryPas encore d'évaluation
- Cyto - Protéine G Et SignalisationDocument30 pagesCyto - Protéine G Et SignalisationOuarib ZakariaPas encore d'évaluation
- Signalisation Par Des Récepteurs TransmembranairesDocument51 pagesSignalisation Par Des Récepteurs TransmembranairesHamza HafidiPas encore d'évaluation
- Récepteurs MembranairesDocument56 pagesRécepteurs MembranairesAnonymous MKSfyYyODPPas encore d'évaluation
- Cours Pharmacodynamie Etudiants2Document92 pagesCours Pharmacodynamie Etudiants2akizar Zinouk100% (1)
- 2.-Biosynthèse Des ProtéinesDocument65 pages2.-Biosynthèse Des Protéinesmido benPas encore d'évaluation
- Cour Régulation Des Gènes Université MentouriDocument16 pagesCour Régulation Des Gènes Université Mentourimonecole2018Pas encore d'évaluation
- Résumé GCDocument4 pagesRésumé GCLilia TlbPas encore d'évaluation
- Correction Devoir de Génétique MoléculaireDocument3 pagesCorrection Devoir de Génétique MoléculairessggnfPas encore d'évaluation
- Métabolisme Du GlycogèneDocument7 pagesMétabolisme Du GlycogèneaxelPas encore d'évaluation
- Eicosanoides PDFDocument5 pagesEicosanoides PDFBouyelfane MohamedPas encore d'évaluation
- Resistance To Thyroid Hormone: SFE Paris 2013 / Annales D'endocrinologie 74 (2013) 238-239Document1 pageResistance To Thyroid Hormone: SFE Paris 2013 / Annales D'endocrinologie 74 (2013) 238-239henry hernandezPas encore d'évaluation
- Httpselearning Centre-Univ-Mila Dza2024pluginfile Php89198mod - resourcecontent1CHAPITRE20V PDFDocument20 pagesHttpselearning Centre-Univ-Mila Dza2024pluginfile Php89198mod - resourcecontent1CHAPITRE20V PDFariel.2mariemPas encore d'évaluation
- Cour Regu 1Document10 pagesCour Regu 1rekik hibaPas encore d'évaluation
- Chapitre 5signalisationDocument7 pagesChapitre 5signalisationFatïma ZohraPas encore d'évaluation
- Cours 2Document39 pagesCours 2fatiPas encore d'évaluation
- Mi To Chon DriesDocument9 pagesMi To Chon Driesamira14.mira83Pas encore d'évaluation
- Régulation Physiologique Des Interactions Entre Hormones Et RécepteursDocument9 pagesRégulation Physiologique Des Interactions Entre Hormones Et RécepteursRyan100% (1)
- Chap II Récepteurs Membranaires Et Molécules de Signalisation IntracellulaireDocument28 pagesChap II Récepteurs Membranaires Et Molécules de Signalisation IntracellulaireAbdoulaye sebyPas encore d'évaluation
- 6 - ProstaglandinesDocument10 pages6 - Prostaglandinesdjamila benaissaPas encore d'évaluation
- BiocellulaireDocument4 pagesBiocellulairecarlio12Pas encore d'évaluation
- Cours - 4 Regulation - Du - Metabolisme - Des - GlucidesDocument18 pagesCours - 4 Regulation - Du - Metabolisme - Des - Glucidesmiss anthropocenePas encore d'évaluation
- Qe BiocellDocument9 pagesQe BiocellASSIA EL MOUSTAID100% (1)
- Endocrino Tableau Récap CC1Document3 pagesEndocrino Tableau Récap CC1BioLab SUPas encore d'évaluation
- 03-Glycolyse 2021Document7 pages03-Glycolyse 2021Mery Sage100% (1)
- Fiche TD Signalisation Cellulaire L3 BPA S6Document7 pagesFiche TD Signalisation Cellulaire L3 BPA S6Massouh Assoui100% (3)
- Communication IntercellulaireDocument5 pagesCommunication Intercellulaireyouness.khalfaouiPas encore d'évaluation
- Chapitre 3 La Régulation de L'expression Des GènesDocument16 pagesChapitre 3 La Régulation de L'expression Des GènesalmnaouarPas encore d'évaluation
- Biochimie1an Glycolyse2022ferganiDocument8 pagesBiochimie1an Glycolyse2022ferganiKhelassi OussamaPas encore d'évaluation
- Le GlucagonDocument6 pagesLe GlucagonRacem Boudghene stambouliPas encore d'évaluation
- Fichier Produit 1945Document64 pagesFichier Produit 1945Antz DanielPas encore d'évaluation
- Chapitre III Récepteurs Et Effets BiologiquesDocument19 pagesChapitre III Récepteurs Et Effets Biologiquesrobert levandoPas encore d'évaluation
- Biochimie2an Glycolyse2019kouiderDocument11 pagesBiochimie2an Glycolyse2019kouiderrahma bouadamPas encore d'évaluation
- Pharmacie 2014 CheminDocument127 pagesPharmacie 2014 Chemindraboissouf174Pas encore d'évaluation
- Cours Chimie Des ColorantsxDocument33 pagesCours Chimie Des Colorantsxdraboissouf174Pas encore d'évaluation
- Feuilletage 2812Document25 pagesFeuilletage 2812draboissouf174Pas encore d'évaluation
- Chapitre1 Biochimie Cellulaire Et FonctionnelleDocument12 pagesChapitre1 Biochimie Cellulaire Et Fonctionnelledraboissouf174Pas encore d'évaluation
- Exercices Sur L'information GénétiqueDocument4 pagesExercices Sur L'information Génétiquedraboissouf174Pas encore d'évaluation
- TD2 Génie GénétiqueDocument2 pagesTD2 Génie Génétiquedraboissouf174Pas encore d'évaluation
- Bumed T 2014 Jeanpierre EmmanuelDocument155 pagesBumed T 2014 Jeanpierre Emmanueldraboissouf174Pas encore d'évaluation
- Cours de Mineralogie2 412Document3 pagesCours de Mineralogie2 412draboissouf174Pas encore d'évaluation
- Examen1 Biochimie MédicaleDocument2 pagesExamen1 Biochimie Médicaledraboissouf174Pas encore d'évaluation
- Ue7 Cours 26Document14 pagesUe7 Cours 26draboissouf174Pas encore d'évaluation
- Katalog Sebia 2015 EngDocument10 pagesKatalog Sebia 2015 EngDarko MaksimovicPas encore d'évaluation
- Corrige Ratrappage BiochiDocument3 pagesCorrige Ratrappage BiochigeovaniePas encore d'évaluation
- Polycopier Génétique Des EucaryotesDocument72 pagesPolycopier Génétique Des EucaryotessabrinaPas encore d'évaluation
- ARN InterférenceDocument3 pagesARN InterférenceshafikPas encore d'évaluation
- La Matrice Extracellulaire 2015-2016 PDFDocument3 pagesLa Matrice Extracellulaire 2015-2016 PDFAnonymous wVr908lBcK100% (2)
- HemostaseDocument8 pagesHemostaseHanane JibetPas encore d'évaluation
- SPS Project 2023Document2 pagesSPS Project 2023ania chichiPas encore d'évaluation
- GénétiqueDocument150 pagesGénétiquefranck brice bado100% (1)
- 1 - Structures Et Propriétés Des Acides NucléiquesDocument19 pages1 - Structures Et Propriétés Des Acides Nucléiquesfousseni karaPas encore d'évaluation
- 07 - Les Rôles de La Membrane PlasmiqueDocument25 pages07 - Les Rôles de La Membrane PlasmiqueSara AllasPas encore d'évaluation
- td2 01Document2 pagestd2 01Lina BiologistaPas encore d'évaluation
- Exploration Des Proteines Plasmatiques 3èmeDocument10 pagesExploration Des Proteines Plasmatiques 3èmeWafa MinamPas encore d'évaluation
- QCM6 La CytosqueletteDocument3 pagesQCM6 La Cytosquelettesarra takfaouiPas encore d'évaluation
- 5-Les Technologies de l'ADN RecombinantDocument7 pages5-Les Technologies de l'ADN RecombinantWi SsalPas encore d'évaluation
- TP Les Fonctions & Les Structures de DonnéesDocument1 pageTP Les Fonctions & Les Structures de Donnéesnasroddine mansourPas encore d'évaluation
- EnzymologieDocument16 pagesEnzymologieFanny GrouxPas encore d'évaluation
- Cours Complet 2022Document34 pagesCours Complet 2022Hhh GbbPas encore d'évaluation
- TD 1 CorrectionDocument13 pagesTD 1 CorrectionBernardine ViouPas encore d'évaluation
- Chapitre 2 La RéplicationDocument18 pagesChapitre 2 La RéplicationHamza Ali100% (1)
- Structure de l'ADNDocument5 pagesStructure de l'ADNWissam TizaPas encore d'évaluation
- Chapitre 7 Et 8 2020Document70 pagesChapitre 7 Et 8 2020Fayol MpayPas encore d'évaluation
- Le Marquage Des Acides NucléiquesDocument5 pagesLe Marquage Des Acides NucléiquesBechir DjazouliPas encore d'évaluation
- Physiologie de L'hémostaseDocument5 pagesPhysiologie de L'hémostaseNouna nouniaPas encore d'évaluation
- Ciam 1SMDocument13 pagesCiam 1SMaboubacarsilecamara92Pas encore d'évaluation
- Chapitre 4 Enzymes-Eleve 2023Document18 pagesChapitre 4 Enzymes-Eleve 2023darwin13jahwin23Pas encore d'évaluation
- TP 5 XerodermaDocument3 pagesTP 5 XerodermaAdam GibaPas encore d'évaluation
- COURS 9 SéquençageDocument109 pagesCOURS 9 SéquençageHikari KazuePas encore d'évaluation
- Bioch1an16-Enzymologie GhoualiDocument67 pagesBioch1an16-Enzymologie Ghoualiahlemm100% (1)
- M Righini Revue BibliographiqueDocument20 pagesM Righini Revue BibliographiqueManon righiniPas encore d'évaluation
- Cours Sv5bcm Genostr1 IzaabelDocument24 pagesCours Sv5bcm Genostr1 Izaabelislmioun100% (1)
- Améliorer votre mémoire: Un Guide pour l'augmentation de la puissance du cerveau, utilisant des techniques et méthodesD'EverandAméliorer votre mémoire: Un Guide pour l'augmentation de la puissance du cerveau, utilisant des techniques et méthodesÉvaluation : 5 sur 5 étoiles5/5 (2)
- Chromatographie: Les Grands Articles d'UniversalisD'EverandChromatographie: Les Grands Articles d'UniversalisPas encore d'évaluation
- Harmonisation Energétique des Lieux: Habitat et haut-lieux sacrés 2020D'EverandHarmonisation Energétique des Lieux: Habitat et haut-lieux sacrés 2020Évaluation : 2.5 sur 5 étoiles2.5/5 (3)
- Le CODE DE DIEU: Le secret de notre passé, la promesse de notre avenirD'EverandLe CODE DE DIEU: Le secret de notre passé, la promesse de notre avenirÉvaluation : 4 sur 5 étoiles4/5 (23)
- Pourquoi suis-je différent?: Accepter et découvrir la puissance de l'hypersensibilitéD'EverandPourquoi suis-je différent?: Accepter et découvrir la puissance de l'hypersensibilitéPas encore d'évaluation
- Les 10 Secrets pour une Vie Plus Heureuse avec la Maladie de ParkinsonD'EverandLes 10 Secrets pour une Vie Plus Heureuse avec la Maladie de ParkinsonPas encore d'évaluation
- Méthode H: Kit de survie scolaire - Apprendre à retenirD'EverandMéthode H: Kit de survie scolaire - Apprendre à retenirPas encore d'évaluation
- La vie des abeilles: Prix Nobel de littératureD'EverandLa vie des abeilles: Prix Nobel de littératureÉvaluation : 4 sur 5 étoiles4/5 (41)
- Guide De Démarrage De L'Apiculture: Le Guide Complet Pour Élever Des AbeillesD'EverandGuide De Démarrage De L'Apiculture: Le Guide Complet Pour Élever Des AbeillesPas encore d'évaluation
- La Conscience Et L'Univers Existent Sans Commencement Ni FinD'EverandLa Conscience Et L'Univers Existent Sans Commencement Ni FinPas encore d'évaluation
- Cahier de jeux de stimulation cognitive: Sujets Alzheimer, désorientés, démences, amnésiesD'EverandCahier de jeux de stimulation cognitive: Sujets Alzheimer, désorientés, démences, amnésiesPas encore d'évaluation
- Nutrition Sportive Fondamentaux et guide pratique du succèsD'EverandNutrition Sportive Fondamentaux et guide pratique du succèsÉvaluation : 5 sur 5 étoiles5/5 (1)
- Géologie de l'Amérique: Les Grands Articles d'UniversalisD'EverandGéologie de l'Amérique: Les Grands Articles d'UniversalisPas encore d'évaluation
- Microbiologie médicale I: agents pathogènes et microbiome humainD'EverandMicrobiologie médicale I: agents pathogènes et microbiome humainÉvaluation : 5 sur 5 étoiles5/5 (1)
- Transition énergétique ET Changement climatique: Développements et Perspectives d'AvenirD'EverandTransition énergétique ET Changement climatique: Développements et Perspectives d'AvenirPas encore d'évaluation
- Histoire de la psychologie scientifique: De la naissance de la psychologie à la neuropsychologie et aux champs d'application les plus actuelsD'EverandHistoire de la psychologie scientifique: De la naissance de la psychologie à la neuropsychologie et aux champs d'application les plus actuelsPas encore d'évaluation
- Appareil génital: Les Grands Articles d'UniversalisD'EverandAppareil génital: Les Grands Articles d'UniversalisPas encore d'évaluation
- Les Graines d'Abricot - Remède de Cancer avec de la Vitamine B17 ?: La Médecine Antique Que l'Industrie Pharmaceutique Moderne CacheD'EverandLes Graines d'Abricot - Remède de Cancer avec de la Vitamine B17 ?: La Médecine Antique Que l'Industrie Pharmaceutique Moderne CachePas encore d'évaluation
- La thérapie libre: La Thérapie de LIBération et Résilience EmotionnelleD'EverandLa thérapie libre: La Thérapie de LIBération et Résilience EmotionnelleÉvaluation : 5 sur 5 étoiles5/5 (1)
- Qu'arrive t'il a la terre arable ?: Le problème de l'érosion du sol.D'EverandQu'arrive t'il a la terre arable ?: Le problème de l'érosion du sol.Évaluation : 5 sur 5 étoiles5/5 (1)
- Macromolécules: Les Grands Articles d'UniversalisD'EverandMacromolécules: Les Grands Articles d'UniversalisPas encore d'évaluation
- Le rêve et ses symboles: Plus de 500 000 exemplaires vendus!D'EverandLe rêve et ses symboles: Plus de 500 000 exemplaires vendus!Évaluation : 5 sur 5 étoiles5/5 (2)
- Atlas permanent de l'Union européenne: 5e édition revue et augmentéeD'EverandAtlas permanent de l'Union européenne: 5e édition revue et augmentéePas encore d'évaluation
- L'élevage professionnel d'insectes: Points stratégiques et méthode de conduiteD'EverandL'élevage professionnel d'insectes: Points stratégiques et méthode de conduitePas encore d'évaluation