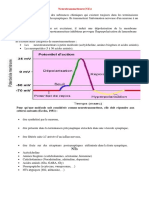
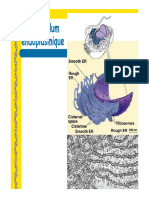
Vous aimerez peut-être aussi
- 11 - Chap11-Les Différents Modes D'action Des HormonesDocument6 pages11 - Chap11-Les Différents Modes D'action Des HormonesYassine HassanePas encore d'évaluation
- Résumé GCDocument4 pagesRésumé GCLilia TlbPas encore d'évaluation
- cibles moléculaires et élaboration de la réponse 2024Document3 pagescibles moléculaires et élaboration de la réponse 2024kaouaneamdjadPas encore d'évaluation
- UE2-Intégration - 13 - Communication-IntercellulaireDocument17 pagesUE2-Intégration - 13 - Communication-Intercellulairesun-nee-chan9Pas encore d'évaluation
- Interaction Ligand RecepteurDocument3 pagesInteraction Ligand RecepteurAmina MnPas encore d'évaluation
- Chapitre 5signalisationDocument7 pagesChapitre 5signalisationFatïma ZohraPas encore d'évaluation
- Définition de La Protéine GDocument4 pagesDéfinition de La Protéine GAsma MhPas encore d'évaluation
- Cyto - Protéine G Et SignalisationDocument30 pagesCyto - Protéine G Et SignalisationOuarib ZakariaPas encore d'évaluation
- RegulationDocument30 pagesRegulationYannick DsprbsPas encore d'évaluation
- Communication CellulaireDocument22 pagesCommunication CellulaireAmïn Fïlalï100% (2)
- Httpselearning Centre-Univ-Mila Dza2024pluginfile Php89198mod - resourcecontent1CHAPITRE20V PDFDocument20 pagesHttpselearning Centre-Univ-Mila Dza2024pluginfile Php89198mod - resourcecontent1CHAPITRE20V PDFariel.2mariemPas encore d'évaluation
- Récepteurs MembranairesDocument56 pagesRécepteurs MembranairesAnonymous MKSfyYyODPPas encore d'évaluation
- Interaction Ligand RécepteurDocument6 pagesInteraction Ligand RécepteurFaculté De Médecine BécharPas encore d'évaluation
- Récepteurs Couplés Au Protéines G Partie 2Document7 pagesRécepteurs Couplés Au Protéines G Partie 2Fatima ZahraPas encore d'évaluation
- Correction Devoir de Génétique MoléculaireDocument3 pagesCorrection Devoir de Génétique MoléculairessggnfPas encore d'évaluation
- Physio1an-Interactions Ligand Recepteur2021bougrida BoussoufDocument24 pagesPhysio1an-Interactions Ligand Recepteur2021bougrida BoussoufGI 2 Session 2018-2019Pas encore d'évaluation
- Neuro 2Document12 pagesNeuro 2tinaPas encore d'évaluation
- La Liaison Aux RécepteursDocument8 pagesLa Liaison Aux RécepteursIsmaelPas encore d'évaluation
- Biologie CellulaireDocument26 pagesBiologie CellulaireBourobou bourobou Chadrack gaelPas encore d'évaluation
- le Signal de Transduction MembranaireDocument25 pagesle Signal de Transduction MembranairekerbabdjamilaPas encore d'évaluation
- Fiche TD Signalisation Cellulaire L3 BPA S6Document7 pagesFiche TD Signalisation Cellulaire L3 BPA S6Massouh Assoui100% (3)
- Hormones CathecholaminesDocument8 pagesHormones Cathecholamineshananehadef04Pas encore d'évaluation
- Interactions Ligand - Recepteur 2023-2024Document29 pagesInteractions Ligand - Recepteur 2023-2024khaldiniheddoc23Pas encore d'évaluation
- Résumé Chapitre 4 Biochimie Cellulaire Et FonctionnelleDocument30 pagesRésumé Chapitre 4 Biochimie Cellulaire Et Fonctionnelledraboissouf174Pas encore d'évaluation
- Ligands Recepteur 2020 EtudiantDocument7 pagesLigands Recepteur 2020 EtudiantLina AmiraPas encore d'évaluation
- Appareil de GolgiDocument4 pagesAppareil de GolgiGericault IfoaratianaPas encore d'évaluation
- Chap II Récepteurs Membranaires Et Molécules de Signalisation IntracellulaireDocument28 pagesChap II Récepteurs Membranaires Et Molécules de Signalisation IntracellulaireAbdoulaye sebyPas encore d'évaluation
- 01 - La Communication IntercellulaireDocument8 pages01 - La Communication IntercellulaireHadil BenslamaPas encore d'évaluation
- Cours de Cytologie, Embryologie Et Genetique Medicale by MBAILASSEM MBAINAIDA GEORGESDocument102 pagesCours de Cytologie, Embryologie Et Genetique Medicale by MBAILASSEM MBAINAIDA GEORGESMãhąmãţ Hãbįb Mõųşşã ĄhmąţPas encore d'évaluation
- Master 2 (Chap.1)Document6 pagesMaster 2 (Chap.1)Ažį ŽaPas encore d'évaluation
- Biologie CellulaireDocument37 pagesBiologie Cellulairenihalaouam05Pas encore d'évaluation
- Le Glutamate Neurotransmetteur: Acide Aminé Synthétisé Par Le Corps Lui-MêmeDocument9 pagesLe Glutamate Neurotransmetteur: Acide Aminé Synthétisé Par Le Corps Lui-MêmeHangouangPas encore d'évaluation
- Les Muscles LissesDocument5 pagesLes Muscles LissesHassanElhassanPas encore d'évaluation
- Cours 4 MaturationDocument42 pagesCours 4 Maturationjeangredin2Pas encore d'évaluation
- 08.10.2015 Organisation Du Trafic IntracellulaireDocument17 pages08.10.2015 Organisation Du Trafic IntracellulaireEaPas encore d'évaluation
- 11 Les Canaux IoniquesDocument12 pages11 Les Canaux IoniquesJean-Loïc BauchetPas encore d'évaluation
- Sujeujet Bio Celle Session 2 PCEM1Document4 pagesSujeujet Bio Celle Session 2 PCEM1medPas encore d'évaluation
- BIO111 ChapitreI Suite 01Document5 pagesBIO111 ChapitreI Suite 01Berthe Aristide Ngue Nkongo100% (1)
- GB3!09!10-Signalisation-Cours E. Macia Généralités Et RCPGDocument118 pagesGB3!09!10-Signalisation-Cours E. Macia Généralités Et RCPGlobnaPas encore d'évaluation
- REF Cytologie PVDocument11 pagesREF Cytologie PVHelena HINTZER--FALCOPas encore d'évaluation
- Les Neurotransmetteurs PacesDocument30 pagesLes Neurotransmetteurs PacesValentin GeorgetPas encore d'évaluation
- METABOLISME CEREBRAL - FggvfelixDocument4 pagesMETABOLISME CEREBRAL - Fggvfelixyoussef majdPas encore d'évaluation
- La Transmission Synaptique 1Document6 pagesLa Transmission Synaptique 1safemindPas encore d'évaluation
- 01 C Interaction Ligand RécepteurDocument39 pages01 C Interaction Ligand RécepteurKhelifi SarraPas encore d'évaluation
- TD7 Les Organites de La Cellule PDFDocument42 pagesTD7 Les Organites de La Cellule PDFZineb DahmaniPas encore d'évaluation
- 14 Histophysiologie Du Tissu NerveuxDocument7 pages14 Histophysiologie Du Tissu NerveuxGeorge BorsPas encore d'évaluation
- Cellules Et Tissus: Objectif GénéralDocument71 pagesCellules Et Tissus: Objectif GénéralDOSSOU EspoirPas encore d'évaluation
- Cours 10 SignalisationDocument21 pagesCours 10 Signalisationshakaraman123Pas encore d'évaluation
- Histamine Et Récepteurs HistalinergiquesDocument12 pagesHistamine Et Récepteurs HistalinergiquesMond DayPas encore d'évaluation
- Chapitre 1Document16 pagesChapitre 1David FMPas encore d'évaluation
- PDF QCM1 Métabolisme-GlucidiqueDocument1 pagePDF QCM1 Métabolisme-GlucidiqueYendouli LAMBONIPas encore d'évaluation
- Réticulum, Golgi Et LysosomesDocument5 pagesRéticulum, Golgi Et LysosomesgeraPas encore d'évaluation
- Compartiments Intracell Et Tri Des ProtéinesDocument43 pagesCompartiments Intracell Et Tri Des Protéinesa29921Pas encore d'évaluation
- DEVOIR DE REGULATION DU METABOLISMEDocument11 pagesDEVOIR DE REGULATION DU METABOLISMEtchabausidoniePas encore d'évaluation
- Cours Pharmacodynamie Etudiants2Document92 pagesCours Pharmacodynamie Etudiants2akizar Zinouk100% (1)
- Métabolisme Du GlycogèneDocument7 pagesMétabolisme Du GlycogèneaxelPas encore d'évaluation
- Pancreas EndocrineDocument4 pagesPancreas Endocrinemalakzerizer2005Pas encore d'évaluation
- 5 La Membrane PlasmiqueDocument11 pages5 La Membrane PlasmiqueKí KŷuśặkuPas encore d'évaluation
- SUPPORT UE1B-KREJBICH-Homéostasie 2-17.10.22Document39 pagesSUPPORT UE1B-KREJBICH-Homéostasie 2-17.10.22Yann legendrePas encore d'évaluation
- Les Vrais Secrets de La Magie NoireDocument113 pagesLes Vrais Secrets de La Magie NoireKawtar Gloom GOthside ൃ100% (1)
- Chapitre5 DHCPDocument17 pagesChapitre5 DHCPjeanPas encore d'évaluation
- Lantschoot, Arnold Van. Un Texte Palimpseste de Vat. Copte 65, Muséon 60 (1947), 261-268Document8 pagesLantschoot, Arnold Van. Un Texte Palimpseste de Vat. Copte 65, Muséon 60 (1947), 261-268yadatanPas encore d'évaluation
- Niveaux Langue Registres Exercices PDFDocument4 pagesNiveaux Langue Registres Exercices PDFSadek Kadda Wahid0% (1)
- Met S 23 Sy PDocument4 pagesMet S 23 Sy Pkhocine100% (1)
- Qui A Inventé L'aiguille CreuseDocument4 pagesQui A Inventé L'aiguille CreusejewelgoodremPas encore d'évaluation
- Manuel D'utilisation Gamme Murale Ar5000 1.1 Samsung 2014Document57 pagesManuel D'utilisation Gamme Murale Ar5000 1.1 Samsung 2014ddrumeaPas encore d'évaluation
- TEXTE N°1: La Jeune AcrobateDocument5 pagesTEXTE N°1: La Jeune Acrobatelouaslama0106Pas encore d'évaluation
- Mécanique Des RochesDocument41 pagesMécanique Des RochesIssam ZougariPas encore d'évaluation
- Les Maladies de Dépérissement Des AgrumesDocument48 pagesLes Maladies de Dépérissement Des AgrumesMichelAndriamahazonoroRaherimanantsoaPas encore d'évaluation
- Bac - Les Meilleures Citations Pour Gagner Des Points en Français - SAMABACDocument4 pagesBac - Les Meilleures Citations Pour Gagner Des Points en Français - SAMABACbeavoguipaulbarre047Pas encore d'évaluation
- 17M110hhhhhh PDFDocument101 pages17M110hhhhhh PDFilham haniniPas encore d'évaluation
- 2-La Hierarchie AngéliqueDocument6 pages2-La Hierarchie Angéliquemichel fayettetchibinda100% (2)
- Les Courants LitterairesDocument2 pagesLes Courants LitterairesAna-Maria RoșuPas encore d'évaluation
- Etude Typologique de L'expression de L'espaceDocument41 pagesEtude Typologique de L'expression de L'espacevj03ePas encore d'évaluation
- Repercussion Islam Ames Hamad1Document15 pagesRepercussion Islam Ames Hamad1saydou bokotaPas encore d'évaluation
- Comptabilité Des CliniquesDocument47 pagesComptabilité Des CliniquesAnonymous kAVA6ALXNPas encore d'évaluation
- TD PalettisationDocument12 pagesTD PalettisationZine eddine Hadj mokhnachrPas encore d'évaluation
- ANALYSE D'UNE Démarche de Cartographie Du RisqueDocument174 pagesANALYSE D'UNE Démarche de Cartographie Du RisqueSouad TouchliftPas encore d'évaluation
- Pied Au PlancherDocument2 pagesPied Au PlancherKenza BenmokhtarPas encore d'évaluation
- Picardie Cerfa 11415-2Document3 pagesPicardie Cerfa 11415-2vanessa.roch1819Pas encore d'évaluation
- Robe Salopette CatherineDocument57 pagesRobe Salopette CatherineAude EPIARD100% (1)
- Corro Philo Valesse 1iere - PDFDocument114 pagesCorro Philo Valesse 1iere - PDFnayoussayacoubou17Pas encore d'évaluation
- Chimie GeneraleDocument5 pagesChimie Generaletcheva jokhanan TiambiPas encore d'évaluation
- Gestion Du Temps KaufDocument44 pagesGestion Du Temps KaufHodaPas encore d'évaluation
- TABLEAU EDEN ROCK - XLSX 2 VILLADocument20 pagesTABLEAU EDEN ROCK - XLSX 2 VILLARebs RebsPas encore d'évaluation
- Rapport Cable TechDocument17 pagesRapport Cable TechMallouki Med100% (1)
- Regarder Le Tresor Du Petit Nicolas Entier VFDocument4 pagesRegarder Le Tresor Du Petit Nicolas Entier VFxzcvxzPas encore d'évaluation
- Wayser2009 2Document25 pagesWayser2009 2ALEXANDRE F VOLTAPas encore d'évaluation
- 3718 v2 Planchers Nervurs Poutrelles Prfa Associes Du Bton Coul en OeuvreDocument98 pages3718 v2 Planchers Nervurs Poutrelles Prfa Associes Du Bton Coul en OeuvreZizouk FeresPas encore d'évaluation