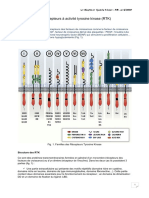
Vous aimerez peut-être aussi
- Maladies NutritionnellesDocument18 pagesMaladies NutritionnellesFatima Zahra83% (6)
- Recepteur A Activite Serine Threonine KinaseDocument11 pagesRecepteur A Activite Serine Threonine KinaseFatima ZahraPas encore d'évaluation
- Bioch S2Document36 pagesBioch S2Eduard SimaderPas encore d'évaluation
- Les Récepteurs À Activité Tyrosine KinaseDocument10 pagesLes Récepteurs À Activité Tyrosine KinaseFatima Zahra100% (1)
- Recepteurs Associes A Une Tyrosine KinaseDocument13 pagesRecepteurs Associes A Une Tyrosine KinaseFatima ZahraPas encore d'évaluation
- SCH Mas Cours SignalisationDocument35 pagesSCH Mas Cours SignalisationWael HahaPas encore d'évaluation
- Les Récepteurs Membranaires Les Récepteurs Couplés Aux Protéines G (RCPG)Document79 pagesLes Récepteurs Membranaires Les Récepteurs Couplés Aux Protéines G (RCPG)Acide OléiquePas encore d'évaluation
- Examen 2017 Normal CorrigeDocument7 pagesExamen 2017 Normal Corrigefawzi badiPas encore d'évaluation
- Chapitre II Expression Des Gènes (Transcription Et Traduction) Leur RégulationDocument9 pagesChapitre II Expression Des Gènes (Transcription Et Traduction) Leur Régulationsolo gamer50% (2)
- Corrigé Exercice PCR 2020Document11 pagesCorrigé Exercice PCR 2020Lara TabetPas encore d'évaluation
- Epissage Chez Les EucaryotesDocument6 pagesEpissage Chez Les EucaryotesAlain Bertrand JuniorPas encore d'évaluation
- 1-Troubles Du Métabolisme Des Glucides PDFDocument5 pages1-Troubles Du Métabolisme Des Glucides PDFAisSa KeRrøùùm100% (1)
- Série 03Document2 pagesSérie 03Moussaoui KhouiraPas encore d'évaluation
- 7 - PCRDocument23 pages7 - PCRVino DongaPas encore d'évaluation
- Cours V La Réaction InflammatoireDocument14 pagesCours V La Réaction InflammatoiretinaPas encore d'évaluation
- Outils de Genie Genetique - 5Document17 pagesOutils de Genie Genetique - 5salama2011Pas encore d'évaluation
- TD Génomique Master UFR Sci Bio DR DagoDocument11 pagesTD Génomique Master UFR Sci Bio DR DagoFANKELE ALASSANE KONE100% (1)
- !!!!!!!!cours Biologie Moleculaie S5Document46 pages!!!!!!!!cours Biologie Moleculaie S5Miro100% (1)
- Genetique Bacterienne 2021Document12 pagesGenetique Bacterienne 2021Oum LoayePas encore d'évaluation
- Contôle Rattrapage Biomol 2021-2022Document4 pagesContôle Rattrapage Biomol 2021-2022IzikoPas encore d'évaluation
- TD Génomique Master UFR Sci Bio DR DagoDocument18 pagesTD Génomique Master UFR Sci Bio DR DagoPrince Otti's Officiel100% (1)
- 1388 Outils de La Biologie Moleculaire M AberkaneDocument86 pages1388 Outils de La Biologie Moleculaire M AberkaneZakariya nadirPas encore d'évaluation
- Les Protozooses 2A Bio 2018Document31 pagesLes Protozooses 2A Bio 2018حمزة بن أحمدPas encore d'évaluation
- Lpro TD 1 Oct16 PDFDocument10 pagesLpro TD 1 Oct16 PDFAnonymous FYcmbifCtmPas encore d'évaluation
- La Spermatogenèse CoursDocument11 pagesLa Spermatogenèse CoursNadjib A. NedjarPas encore d'évaluation
- Solution Exercise I:: Soit La Séquence Suivante Correspondant Au Gène X de Sinorhizibium MelilotiDocument2 pagesSolution Exercise I:: Soit La Séquence Suivante Correspondant Au Gène X de Sinorhizibium MelilotiITAW100% (1)
- Exercice S 2016Document17 pagesExercice S 2016Aboubakar LawanePas encore d'évaluation
- Séquençage de l'ADN PDFDocument41 pagesSéquençage de l'ADN PDFChibani ZinebPas encore d'évaluation
- Tri Proteine 2014 15Document131 pagesTri Proteine 2014 15chaimaePas encore d'évaluation
- Td2 Corrige TypeDocument3 pagesTd2 Corrige TypeAsmae EssarhiPas encore d'évaluation
- Génétique Microbienne PDFDocument65 pagesGénétique Microbienne PDFAnonymous MKSfyYyODPPas encore d'évaluation
- MyelomeDocument59 pagesMyelomeNardjes DjedouaniPas encore d'évaluation
- $TD #1 D O.B PDFDocument2 pages$TD #1 D O.B PDFNacera CeraPas encore d'évaluation
- Le Systeme Du ComplementDocument8 pagesLe Systeme Du ComplementAymenPas encore d'évaluation
- Génie GenetiqueDocument36 pagesGénie GenetiqueMoussaoui Khouira100% (1)
- Genetique1an16-Regulation Expression GenesDocument8 pagesGenetique1an16-Regulation Expression GenesAmine BeninePas encore d'évaluation
- Corrige Applications Genie Genetique L3 BI Section A Et B 2020 2021 - 1 PDFDocument30 pagesCorrige Applications Genie Genetique L3 BI Section A Et B 2020 2021 - 1 PDFNourPas encore d'évaluation
- Proteines 22-23Document52 pagesProteines 22-23Linda Mansour100% (1)
- Devoir de Contrôle n3 2010 2011said Mounir1mhamdiaDocument9 pagesDevoir de Contrôle n3 2010 2011said Mounir1mhamdiaChokri BesbesPas encore d'évaluation
- TD Les Transferts GénétiqueDocument4 pagesTD Les Transferts Génétiquescience علوم100% (1)
- PCRDocument14 pagesPCRopenlabunistraPas encore d'évaluation
- 04-Interaction Protéine LigandsDocument10 pages04-Interaction Protéine LigandsKhadidja SellaiPas encore d'évaluation
- TranscriptionDocument118 pagesTranscriptionanon_473255111100% (1)
- Fiche II TD BIO 111 2020-2021Document2 pagesFiche II TD BIO 111 2020-2021Pius NdjebetPas encore d'évaluation
- Gène Génétique BoucheffaDocument44 pagesGène Génétique Boucheffary yukPas encore d'évaluation
- TD2 Biologie MoléculaireDocument7 pagesTD2 Biologie Moléculairejebranesanaa92Pas encore d'évaluation
- La RT PCRDocument12 pagesLa RT PCRMbacké0% (1)
- Diapos Ed9 ProteomiqueDocument55 pagesDiapos Ed9 ProteomiqueEmna El Hammi100% (1)
- La PCR PDFDocument27 pagesLa PCR PDFfaroukPas encore d'évaluation
- TD Biologie Moléculaire BPR 2021Document21 pagesTD Biologie Moléculaire BPR 2021rafaramalala1307Pas encore d'évaluation
- Chapitre I: L'expression de L'information GénétiqueDocument74 pagesChapitre I: L'expression de L'information Génétiqueahméd_facebPas encore d'évaluation
- Chapitre 1-Les Divisions Cellulaires IMPORTANTDocument2 pagesChapitre 1-Les Divisions Cellulaires IMPORTANTphilippePas encore d'évaluation
- TD3 BV T.conducteur IDocument6 pagesTD3 BV T.conducteur IRahma LionnePas encore d'évaluation
- TP 1mitoseMeioseFSAAM20Document6 pagesTP 1mitoseMeioseFSAAM20IMADPas encore d'évaluation
- Sujet GénétiqueDocument2 pagesSujet GénétiqueM'bouillé SissokoPas encore d'évaluation
- Genetique s5 TD 5Document60 pagesGenetique s5 TD 5dark kristalPas encore d'évaluation
- TD3 Et Corrigé Type 2022Document7 pagesTD3 Et Corrigé Type 2022ines LydiaPas encore d'évaluation
- Examen Génétique S5 JANVIER 2020 VARIANTE 1 Correction DefDocument4 pagesExamen Génétique S5 JANVIER 2020 VARIANTE 1 Correction DeflglawiadikPas encore d'évaluation
- Groupe2 Composes ParietauxDocument32 pagesGroupe2 Composes ParietauxCheick ahmedPas encore d'évaluation
- SVT ResuméDocument18 pagesSVT ResuméAya RyadPas encore d'évaluation
- Cours P3chap3 PDFDocument3 pagesCours P3chap3 PDFhilka féréole mbadingaPas encore d'évaluation
- Cours II Les Déficits Immunitaires Innés - CopieDocument10 pagesCours II Les Déficits Immunitaires Innés - CopietinaPas encore d'évaluation
- Effet Du Stress AbiotiqueDocument96 pagesEffet Du Stress AbiotiqueFatima ZahraPas encore d'évaluation
- Pharmacocinetique Et Toxicologie Des PolluantsDocument14 pagesPharmacocinetique Et Toxicologie Des PolluantsFatima ZahraPas encore d'évaluation
- NFK BDocument8 pagesNFK BFatima ZahraPas encore d'évaluation
- Cours 1 PHYSIOLOGIE DE LA NUTRITION SVI S6 EnseignantDocument32 pagesCours 1 PHYSIOLOGIE DE LA NUTRITION SVI S6 EnseignantFatima Zahra80% (5)
- Jak and StatDocument16 pagesJak and StatFatima ZahraPas encore d'évaluation
- Cours ABSORPTION - PHARMACOCINETIQUE ET TOXICOLOGIE DES POLLUANTSDocument47 pagesCours ABSORPTION - PHARMACOCINETIQUE ET TOXICOLOGIE DES POLLUANTSFatima ZahraPas encore d'évaluation
- Récepteur de La Superfamille TNF AlphaDocument28 pagesRécepteur de La Superfamille TNF AlphaFatima ZahraPas encore d'évaluation
- Récepteurs À Activité Tyrosine Kinase DefDocument12 pagesRécepteurs À Activité Tyrosine Kinase DefFatima ZahraPas encore d'évaluation
- Récepteurs Couplés Au Protéines G Partie 2Document7 pagesRécepteurs Couplés Au Protéines G Partie 2Fatima ZahraPas encore d'évaluation
- Récépteurs Couplés Aux Protéines G ModifDocument22 pagesRécépteurs Couplés Aux Protéines G ModifFatima ZahraPas encore d'évaluation
- M Co Fi 9 Plaquette - Technique - Standard - Histologie PDFDocument2 pagesM Co Fi 9 Plaquette - Technique - Standard - Histologie PDFKhawla ZidouriPas encore d'évaluation
- Immuno3an Lymphocytes-TDocument23 pagesImmuno3an Lymphocytes-TSaid AkcentPas encore d'évaluation
- Mécanismes de La Catalyse Et Action Des EffecteursDocument23 pagesMécanismes de La Catalyse Et Action Des EffecteursRan OuchPas encore d'évaluation
- Physiologie Du Globule Rouge Et Physiopathologie Des AnémiesDocument87 pagesPhysiologie Du Globule Rouge Et Physiopathologie Des AnémiesChahine BlPas encore d'évaluation
- La Découverte de La Molécule ADN - CorrectionDocument3 pagesLa Découverte de La Molécule ADN - CorrectionBeshta AhmadiPas encore d'évaluation
- Examen Biologie Cellulaire 2015+corriger Université Amar Telidji de LaghouatDocument4 pagesExamen Biologie Cellulaire 2015+corriger Université Amar Telidji de Laghouatazzam100% (1)
- Hybridation MoléculaireDocument2 pagesHybridation MoléculaireHÄ ÝÄŤ100% (1)
- Référentiel Virologie 2020-21 UP7 PDFDocument255 pagesRéférentiel Virologie 2020-21 UP7 PDFamyPas encore d'évaluation
- Concours Maitre Assistant 2015Document57 pagesConcours Maitre Assistant 2015Sarra KlaiPas encore d'évaluation
- Biologie Moléculaire Coindre 2010Document9 pagesBiologie Moléculaire Coindre 2010abidPas encore d'évaluation
- Taalim Pro - La Biologie Cellulaire - 2023 - CompressedDocument68 pagesTaalim Pro - La Biologie Cellulaire - 2023 - Compressedhoucine hamdonPas encore d'évaluation
- Cours Immunologie PDFDocument63 pagesCours Immunologie PDFfafi Ali100% (2)
- Chapitre III Les ProtéinesDocument13 pagesChapitre III Les ProtéinesEssassi AmmarPas encore d'évaluation
- Métabolisme Des Acides Aminés Cours de Biochimie 2eme Année Pharmacie DR CHERIETDocument15 pagesMétabolisme Des Acides Aminés Cours de Biochimie 2eme Année Pharmacie DR CHERIETHea FPas encore d'évaluation
- 1.3.01.1ecrit Cours Physiologie Et Exploration Biologique L'hemostase Primaire Et Coagulation++Document36 pages1.3.01.1ecrit Cours Physiologie Et Exploration Biologique L'hemostase Primaire Et Coagulation++malick SemourPas encore d'évaluation
- 1spé T1 Chapitre3 CoursDocument8 pages1spé T1 Chapitre3 Coursabdirahman abdillahiPas encore d'évaluation
- Structure Des BactériesDocument9 pagesStructure Des BactérieskhouildiPas encore d'évaluation
- TD Peptitides Et Protéines 2000-2001Document3 pagesTD Peptitides Et Protéines 2000-2001Ikram FerPas encore d'évaluation
- Cours Proteine 03-12-2Document20 pagesCours Proteine 03-12-2Mohamed Adnen Hadj SessiPas encore d'évaluation
- TD Chapitre 3 Bio Cell 1Document13 pagesTD Chapitre 3 Bio Cell 1satisfying541Pas encore d'évaluation
- Cours de Structure 3D Des Macromolécules Biologiques L3Biomol DERKAOUIDocument241 pagesCours de Structure 3D Des Macromolécules Biologiques L3Biomol DERKAOUIj64491284Pas encore d'évaluation
- UE2-Biologie Cellulaire-Cours 1-Généralités Sur La cellule-1718-FINALDocument10 pagesUE2-Biologie Cellulaire-Cours 1-Généralités Sur La cellule-1718-FINALsmash.00-ragasPas encore d'évaluation
- Analyse Multivariée Des Relations Structures-Activités Dans Des Nouveaux Composés Organiques À Utilisation Médicinale SOUALMIA FatimaDocument130 pagesAnalyse Multivariée Des Relations Structures-Activités Dans Des Nouveaux Composés Organiques À Utilisation Médicinale SOUALMIA Fatimaazou korbaPas encore d'évaluation
- Les Auto Antigènes Et Auto AnticorpsDocument21 pagesLes Auto Antigènes Et Auto AnticorpsSidiraos Ewoneraos Sidi SidiPas encore d'évaluation
- Introduction Biologie Cellulaire 2020Document27 pagesIntroduction Biologie Cellulaire 2020Sani Amadou El-ousmanePas encore d'évaluation
- These Benabdelkader Tarek 2012Document281 pagesThese Benabdelkader Tarek 2012Kima MadPas encore d'évaluation
- Manual Nutricion Dietetica CARBAJALDocument71 pagesManual Nutricion Dietetica CARBAJALAstrid OlartePas encore d'évaluation
- Copie S1BC FMPC22Document10 pagesCopie S1BC FMPC22ilhamdandi523Pas encore d'évaluation