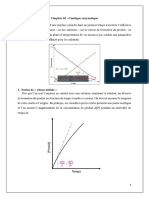
Vous aimerez peut-être aussi
- E97p308 PDFDocument6 pagesE97p308 PDFEL Omrani SaidPas encore d'évaluation
- DMB Une Nageoire Épineuse À Double Tranchant...Document6 pagesDMB Une Nageoire Épineuse À Double Tranchant...JB LimonierPas encore d'évaluation
- Cinétique Enzymatique À 2 SubstratsDocument7 pagesCinétique Enzymatique À 2 Substratsamina44Pas encore d'évaluation
- Cours M2 PINGPONGDocument8 pagesCours M2 PINGPONGAllah Beurem BonheurPas encore d'évaluation
- 06-Cinétique Enzymatique à Deux SubstratsDocument8 pages06-Cinétique Enzymatique à Deux SubstratsSlimkhane DzPas encore d'évaluation
- Cours de Cinetique A Deux Substrats PDFDocument7 pagesCours de Cinetique A Deux Substrats PDFJalelo AbdelPas encore d'évaluation
- ENZYMOLOGIE II.2021pptDocument59 pagesENZYMOLOGIE II.2021pptKaren DominiquePas encore d'évaluation
- Cours de Cinetique A Deux SubstratsDocument8 pagesCours de Cinetique A Deux Substratsmistic JayPas encore d'évaluation
- Materiaux 2-Diag PhasesDocument12 pagesMateriaux 2-Diag PhasesCassie TientcheuPas encore d'évaluation
- Diagrammes Binaires: 2ième PartieDocument69 pagesDiagrammes Binaires: 2ième PartiezemoussaPas encore d'évaluation
- Kira ADocument5 pagesKira AManon righiniPas encore d'évaluation
- Corrige Serie3 PDFDocument5 pagesCorrige Serie3 PDFMohamedDokse0% (1)
- 05extrait Thermo MateriauxDocument69 pages05extrait Thermo MateriauxazuutgPas encore d'évaluation
- Diagramme de PhaseDocument8 pagesDiagramme de PhaseAymen dzPas encore d'évaluation
- 1°sDocument42 pages1°sla physique selon le programme Français100% (2)
- Biochimie1an-Cinetique Enzymatique2020feraga PDFDocument4 pagesBiochimie1an-Cinetique Enzymatique2020feraga PDFAsmaPas encore d'évaluation
- Sujet+correction Ue1 2019Document39 pagesSujet+correction Ue1 2019Nany LovePas encore d'évaluation
- Aproximative Des Laison FortesDocument19 pagesAproximative Des Laison Fortesmohamed mostefaiPas encore d'évaluation
- 18 Acide-Base 1Document3 pages18 Acide-Base 1Ilyes Ben JemaaPas encore d'évaluation
- Séance VI - COURS. Enzymologie UE-SF 2Document51 pagesSéance VI - COURS. Enzymologie UE-SF 2Lamisse AbedPas encore d'évaluation
- EMD 1 Chimie 2013Document2 pagesEMD 1 Chimie 2013Mbaye AbouPas encore d'évaluation
- Les Diagrammes de Phases BinairesDocument69 pagesLes Diagrammes de Phases BinairesYami le Chimiste100% (6)
- Chapitre 3 Diagrammes Equilibre BinaireDocument15 pagesChapitre 3 Diagrammes Equilibre BinaireYoussef GharianiPas encore d'évaluation
- CompDéfini MG ZNDocument4 pagesCompDéfini MG ZNChikov ÆminePas encore d'évaluation
- 2 - Introduction A L - Enzymologie PDFDocument9 pages2 - Introduction A L - Enzymologie PDFGabrielle NnomoPas encore d'évaluation
- Chapitre IV équilibre Liquide-SolideDocument8 pagesChapitre IV équilibre Liquide-SolideChamssou BoustilaPas encore d'évaluation
- Corrigé Type1 PC 1 D .Essai Blanc2024Document5 pagesCorrigé Type1 PC 1 D .Essai Blanc2024issaradjab13Pas encore d'évaluation
- PARTIE C-La DistillationDocument13 pagesPARTIE C-La DistillationSABAPas encore d'évaluation
- Huckel PDFDocument13 pagesHuckel PDFSalah LáálámPas encore d'évaluation
- Chapitre 4 Cinétique Et InhibitionDocument14 pagesChapitre 4 Cinétique Et Inhibitionkgu yryrtyPas encore d'évaluation
- Chimie MecanismesReactionnelsDocument13 pagesChimie MecanismesReactionnelsHmemaPas encore d'évaluation
- HybridationDocument11 pagesHybridationYassine SabekPas encore d'évaluation
- D4 TS1 2023 LVN - WahabdiopDocument4 pagesD4 TS1 2023 LVN - Wahabdiopsokhnamaimounadiop662004Pas encore d'évaluation
- Chapitre 3 Cours Diagrammes D - ÉquilibreDocument21 pagesChapitre 3 Cours Diagrammes D - ÉquilibrebenchallalPas encore d'évaluation
- Equilibres Solide Liquide PDFDocument5 pagesEquilibres Solide Liquide PDFWael ZidPas encore d'évaluation
- Cin Tique Et Catalyse HomogŠnePDocument20 pagesCin Tique Et Catalyse HomogŠnePKhaled BouchaourPas encore d'évaluation
- Algèbre de BooleDocument28 pagesAlgèbre de BooleSaad Ben Belmir100% (1)
- Les Equations de MaxwellDocument24 pagesLes Equations de MaxwellMaria SeltanaPas encore d'évaluation
- 3a - Cinetique EnzymatiqueDocument8 pages3a - Cinetique EnzymatiqueSameh Khemir100% (2)
- 7 - Mécanisme de La CatalyseDocument34 pages7 - Mécanisme de La Catalyseangelo.terrisse1Pas encore d'évaluation
- Diple RLDocument5 pagesDiple RLEmna TräadPas encore d'évaluation
- Cours Therm Chapitre Vi Equilibre Liquide Solide l3 2019 13 AvrilDocument60 pagesCours Therm Chapitre Vi Equilibre Liquide Solide l3 2019 13 AvrilHanane faByyPas encore d'évaluation
- Introduction A La Chimie Moleculaire-Chap4Document8 pagesIntroduction A La Chimie Moleculaire-Chap4Rodolphe BledPas encore d'évaluation
- Séance de QCMs - BiochimieDocument39 pagesSéance de QCMs - Biochimieشكوب ستانPas encore d'évaluation
- 1 Binaires RDocument100 pages1 Binaires ROuday Chikhaoui100% (1)
- Chapitre 02 Cinétique EnzymatiqueDocument5 pagesChapitre 02 Cinétique Enzymatiqueamina44Pas encore d'évaluation
- La Cinétique EnzymatiqueDocument6 pagesLa Cinétique EnzymatiqueTCHEMMIK NazihaPas encore d'évaluation
- Equilibre Solide Liquide OUII Du 1ère masterGCDocument8 pagesEquilibre Solide Liquide OUII Du 1ère masterGCRay CharlesPas encore d'évaluation
- Complement Thevenin NortonDocument4 pagesComplement Thevenin NortonDocteur Albert TouatiPas encore d'évaluation
- Énoncé TPDocument10 pagesÉnoncé TPLemjid YesminePas encore d'évaluation
- Tableau de KarnaughDocument8 pagesTableau de KarnaughJaona RakotonirinaPas encore d'évaluation
- Sujet 5Document6 pagesSujet 5Jihed HorchaniPas encore d'évaluation
- DS 2 09 10 4M T SC - ExpSC PDFDocument7 pagesDS 2 09 10 4M T SC - ExpSC PDFnawalsaadPas encore d'évaluation
- Devoir de Synthèse N°2 - Sciences Physiques - Bac Sciences Exp (2022-2023) MR BARHOUMI EZZEDINEDocument6 pagesDevoir de Synthèse N°2 - Sciences Physiques - Bac Sciences Exp (2022-2023) MR BARHOUMI EZZEDINEGhada DaddouPas encore d'évaluation
- KedDocument3 pagesKedJohn LocationPas encore d'évaluation
- TD 01 EnzymologieDocument2 pagesTD 01 EnzymologieBENYAHIA NESRINEPas encore d'évaluation
- DC2 Q M Copie FINALEDocument4 pagesDC2 Q M Copie FINALEomar sellemi0% (2)
- Exercices d'intégrales et d'équations intégro-différentiellesD'EverandExercices d'intégrales et d'équations intégro-différentiellesPas encore d'évaluation
- Immunologie Fondamentale Partiel 2016Document9 pagesImmunologie Fondamentale Partiel 2016Joe MehannaPas encore d'évaluation
- 101-NYA-05 Etape1 Ch. 7 A23 EA FinalDocument38 pages101-NYA-05 Etape1 Ch. 7 A23 EA FinalFiset st cyr CharliePas encore d'évaluation
- II.1 LE PHYTOCHROME_Photomorphogénèse_Module Croissance et Développement_S5_2022-23Document49 pagesII.1 LE PHYTOCHROME_Photomorphogénèse_Module Croissance et Développement_S5_2022-23Kaoutar OuboualiPas encore d'évaluation
- La Conjugaison 2 - Copie - Etudts - CopieDocument8 pagesLa Conjugaison 2 - Copie - Etudts - CopieMemoire 2223Pas encore d'évaluation
- 1-Généralités Sur Les CellulesDocument15 pages1-Généralités Sur Les Cellulesjacobs100% (1)
- Biotecnologia FRADocument125 pagesBiotecnologia FRAfayçalPas encore d'évaluation
- Génie-Génétique Intro PDFDocument38 pagesGénie-Génétique Intro PDFMaï SsaPas encore d'évaluation
- Questionnaire Bio 2023Document46 pagesQuestionnaire Bio 2023mpogazi clery arecPas encore d'évaluation
- Pharm2an16 Genetique TranscriptionDocument21 pagesPharm2an16 Genetique TranscriptionRadjaa AddPas encore d'évaluation
- Biochimie1an-Cinetique Enzymatique2020feragaDocument27 pagesBiochimie1an-Cinetique Enzymatique2020feragaImene Aoun SeghirPas encore d'évaluation
- Snv-Biologie Cellulaires-Introduction La CelluleDocument3 pagesSnv-Biologie Cellulaires-Introduction La CelluleJoujou HadjinePas encore d'évaluation
- QCM MicrobiologieDocument4 pagesQCM Microbiologiesc biologique80% (10)
- Feuilletage PDFDocument28 pagesFeuilletage PDFAbderrahim BoudaniPas encore d'évaluation
- Les Cellules Eucaryotes 2020 - 2021Document5 pagesLes Cellules Eucaryotes 2020 - 2021Jimingot jams0% (1)
- Cours SVT Tle D&C 2013-2014 Zoure-1Document201 pagesCours SVT Tle D&C 2013-2014 Zoure-1Leonard NGASSAPas encore d'évaluation
- 09 AFLPsDocument26 pages09 AFLPsAzzedine BakaPas encore d'évaluation
- TD 1 BMGG 2024Document4 pagesTD 1 BMGG 2024Abbou BelkaisPas encore d'évaluation
- Les Protéines Du LaitDocument1 pageLes Protéines Du LaitAbu AquiPas encore d'évaluation
- Les Acides Nucléiques 1Document3 pagesLes Acides Nucléiques 1Aymane KrachiPas encore d'évaluation
- Dossier Enseignant Extraction ADNDocument12 pagesDossier Enseignant Extraction ADNCaroline EDU-TAYEBICPas encore d'évaluation
- Cours 5 Clonage Et Vecteurs de ClonageDocument37 pagesCours 5 Clonage Et Vecteurs de ClonageOwassiPas encore d'évaluation
- FORMATION-Laboratoire Techniques PDFDocument38 pagesFORMATION-Laboratoire Techniques PDFAadil Aadil0% (1)
- activité 3 ADN TPDocument2 pagesactivité 3 ADN TPefnt2023Pas encore d'évaluation
- Ram 5159Document127 pagesRam 5159achraf rabadiPas encore d'évaluation
- Cinetique Enzymatique-1 PDFDocument15 pagesCinetique Enzymatique-1 PDFTCHEMMIK NazihaPas encore d'évaluation
- Gnomique ProteomiqueDocument52 pagesGnomique ProteomiqueAsma PrettyangelPas encore d'évaluation
- Anomalies Constitutionnelles de La Coagulation Prédisposant À La Thrombose 2007 EMCDocument18 pagesAnomalies Constitutionnelles de La Coagulation Prédisposant À La Thrombose 2007 EMCJankis KhanPas encore d'évaluation
- Phos CellDocument15 pagesPhos CellAhmed Rabii0% (1)
- 44957349devoir de Controle N 3 Lycee Pilote SVT 2eme Sciences 2009 2010 Mme Abdelmoula Nee Jmal Amel PDFDocument2 pages44957349devoir de Controle N 3 Lycee Pilote SVT 2eme Sciences 2009 2010 Mme Abdelmoula Nee Jmal Amel PDFMohamed Mahfoudh WewahPas encore d'évaluation