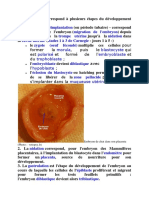
Vous aimerez peut-être aussi
- Appareil génital: Les Grands Articles d'UniversalisD'EverandAppareil génital: Les Grands Articles d'UniversalisPas encore d'évaluation
- 647m1 EMBRYOLOGIEDocument20 pages647m1 EMBRYOLOGIEKamilia BilalPas encore d'évaluation
- EmbryogénèseDocument5 pagesEmbryogénèseredPas encore d'évaluation
- Cours Biologie Du Développement L3 EntomologieDocument48 pagesCours Biologie Du Développement L3 EntomologieRofaida IlmainPas encore d'évaluation
- Introduction Générale Cours - Biologie - Animale-Snv-L1 PDFDocument4 pagesIntroduction Générale Cours - Biologie - Animale-Snv-L1 PDFNabil holmesPas encore d'évaluation
- Cours Biologie Animale SNV l1 2015 Introduction GeneraleDocument4 pagesCours Biologie Animale SNV l1 2015 Introduction GeneraleSabrin Amani100% (1)
- Cours SVT Inter TC SC Reproduction Végétaux Sans FleursDocument28 pagesCours SVT Inter TC SC Reproduction Végétaux Sans FleursTahiri Mehdi100% (2)
- Développement végétal: Les Grands Articles d'UniversalisD'EverandDéveloppement végétal: Les Grands Articles d'UniversalisPas encore d'évaluation
- ChapI Annexes Emb Controle DevDocument6 pagesChapI Annexes Emb Controle DevBabel BodjonaPas encore d'évaluation
- Développement EmbryonnaireDocument9 pagesDéveloppement EmbryonnaireEva Peignier100% (1)
- Chapitre V Gastrulation (Cours)Document2 pagesChapitre V Gastrulation (Cours)jsaadoudanbakoPas encore d'évaluation
- Chapitre VReproduction 114Document83 pagesChapitre VReproduction 114mouhamadoulaminembengue64Pas encore d'évaluation
- Embryologie S1 2014 2015 1Document27 pagesEmbryologie S1 2014 2015 1L'enfant SucréPas encore d'évaluation
- Cours Presentation Bid 2022 l1 SN'Document387 pagesCours Presentation Bid 2022 l1 SN'yssoufPas encore d'évaluation
- Biologie Animale 3eme Trimestre PDFDocument29 pagesBiologie Animale 3eme Trimestre PDFFafi AliPas encore d'évaluation
- Embryo LogieDocument12 pagesEmbryo LogieinthemoodforlifePas encore d'évaluation
- Embryologie S1 - 2020-2021 PolyDocument29 pagesEmbryologie S1 - 2020-2021 PolyDec30mdi BtbPas encore d'évaluation
- Introduction A L'embryologieDocument3 pagesIntroduction A L'embryologieMaster DZPas encore d'évaluation
- Procréation: Les Grands Articles d'UniversalisD'EverandProcréation: Les Grands Articles d'UniversalisPas encore d'évaluation
- Chapitre II Les Mouvements Morphogeniques Dans Lorganisation Spatiale de LembryonDocument24 pagesChapitre II Les Mouvements Morphogeniques Dans Lorganisation Spatiale de LembryonŽdn ÂśšiaPas encore d'évaluation
- Regulation Et Determination Des Cellules EmbryonnairesDocument60 pagesRegulation Et Determination Des Cellules EmbryonnairesGhiles Anis OuyahiaPas encore d'évaluation
- Chapitre 11 Reproduction Des Oiseaux Embryologie A1 Djeffal PDFDocument14 pagesChapitre 11 Reproduction Des Oiseaux Embryologie A1 Djeffal PDFVéto SamPas encore d'évaluation
- CafDocument86 pagesCafLàbÿôuï ÃbderráhmàñëPas encore d'évaluation
- Chapitre I, II, IIIDocument34 pagesChapitre I, II, IIIRodolphe YoboPas encore d'évaluation
- EmbriogeneseDocument16 pagesEmbriogeneseYahiaab RahmaniPas encore d'évaluation
- Protozoaires - MétazoairesDocument39 pagesProtozoaires - MétazoairesdetobPas encore d'évaluation
- Id 3457Document29 pagesId 3457salah a9aPas encore d'évaluation
- Developpe MentDocument71 pagesDeveloppe MentKhalid Jbilou100% (1)
- Revisions Du Cours SVI M2 S1 Avec QCEMDocument52 pagesRevisions Du Cours SVI M2 S1 Avec QCEMazzamPas encore d'évaluation
- ChapI Embryo ExpDocument10 pagesChapI Embryo ExpBabel BodjonaPas encore d'évaluation
- Chapitre 14Document28 pagesChapitre 14TML7Pas encore d'évaluation
- DE_DPE_amphibienDocument26 pagesDE_DPE_amphibienpascalbessan1998Pas encore d'évaluation
- PR Sahraoui La GestationDocument3 pagesPR Sahraoui La Gestation4cprqpbhw5Pas encore d'évaluation
- La SegmentationDocument18 pagesLa SegmentationNabil holmesPas encore d'évaluation
- Le ClonageDocument16 pagesLe Clonageraslenbouallegue12345Pas encore d'évaluation
- GESTATIODocument12 pagesGESTATIOFaustin Rassembaye AllarabayePas encore d'évaluation
- Cours 3 M.aknoun SailDocument9 pagesCours 3 M.aknoun Sailabderrahmanecherchour510Pas encore d'évaluation
- Regne AnimalDocument21 pagesRegne Animalghislainqicoci74Pas encore d'évaluation
- Résumé L2 Agro BotaniqueDocument18 pagesRésumé L2 Agro BotaniqueImane Zayn100% (1)
- Résumé L2 Agro BotaniqueDocument18 pagesRésumé L2 Agro BotaniqueJosh HPas encore d'évaluation
- Développement Embryonnaire de La DrosophileDocument25 pagesDéveloppement Embryonnaire de La DrosophileSarah OlivierPas encore d'évaluation
- Cours 1master BioDevDocument58 pagesCours 1master BioDevRodolphe YoboPas encore d'évaluation
- Chapitre 8 Annexes Embryonnaires-FinalDocument9 pagesChapitre 8 Annexes Embryonnaires-FinalVéto SamPas encore d'évaluation
- ReproductionDocument11 pagesReproductionSamir BenmouffokPas encore d'évaluation
- 4 5 BV 7Document87 pages4 5 BV 7Mohammed EcharradiPas encore d'évaluation
- 0 Chap 1 TH 1A Cours PadletDocument11 pages0 Chap 1 TH 1A Cours Padlet2jm6tpbjfwPas encore d'évaluation
- La Fecondation: Classe:3 C ProfesseurDocument6 pagesLa Fecondation: Classe:3 C Professeurepiphane diogoye mbenguePas encore d'évaluation
- Wa0012Document8 pagesWa0012Babacar SowPas encore d'évaluation
- Cour Biologie Animale Complet (Tarek Hajji) (Isbst)Document54 pagesCour Biologie Animale Complet (Tarek Hajji) (Isbst)Fahroudine100% (1)
- Cours BA Invertébrés SefriouiDocument215 pagesCours BA Invertébrés SefriouiAboulhouda HatubPas encore d'évaluation
- Cours de Zoologie L2Document38 pagesCours de Zoologie L2Hamza KadaPas encore d'évaluation
- (Former La, Puis Le Qui Est Et Formé de L' Et Du Avec L' Et L' Pour Pouvoir S'implanter Dans LaDocument2 pages(Former La, Puis Le Qui Est Et Formé de L' Et Du Avec L' Et L' Pour Pouvoir S'implanter Dans LaRihem BarketPas encore d'évaluation
- Embryologie s1Document127 pagesEmbryologie s1MeRyem El MejdoubPas encore d'évaluation
- PrésentationDocument9 pagesPrésentationZikoox TnPas encore d'évaluation
- ProtozoairesDocument11 pagesProtozoairesBejen BEJENPas encore d'évaluation
- Resume Protozoaires 2Document4 pagesResume Protozoaires 2Maderno manaPas encore d'évaluation
- chap IV la segmentation (support du cours)Document3 pageschap IV la segmentation (support du cours)Sah RkmPas encore d'évaluation
- Faculté de Médecine de Sétif - Algérie Année Universitaire 2019-2020Document23 pagesFaculté de Médecine de Sétif - Algérie Année Universitaire 2019-2020Tholet-man SylvestrePas encore d'évaluation
- COURS DU PROF DAGNOGO PDF+Document56 pagesCOURS DU PROF DAGNOGO PDF+Henri Jowel100% (1)
- Chapitre 4 Segmentation Alouani Cour 2017Document8 pagesChapitre 4 Segmentation Alouani Cour 2017chahinda lakroutPas encore d'évaluation
- TP-TD SolDocument5 pagesTP-TD SolAYDEN GreenPas encore d'évaluation
- Comptabilite Generale Et Analitique D'exploitationDocument27 pagesComptabilite Generale Et Analitique D'exploitationAYDEN GreenPas encore d'évaluation
- Cours BC Chap Iii Et IvDocument19 pagesCours BC Chap Iii Et IvAYDEN GreenPas encore d'évaluation
- Chap3.Deuxième PrincipeDocument8 pagesChap3.Deuxième PrincipeAYDEN GreenPas encore d'évaluation
- Chap5.Représentation Des Évolutions D'un FluideDocument6 pagesChap5.Représentation Des Évolutions D'un FluideAYDEN GreenPas encore d'évaluation
- ELECTROCINETIQUEDocument17 pagesELECTROCINETIQUEAYDEN GreenPas encore d'évaluation
- Chapitre 1:: ElectrostatiqueDocument11 pagesChapitre 1:: ElectrostatiqueAYDEN GreenPas encore d'évaluation
- Fiche VascularisationDocument10 pagesFiche VascularisationAlaa MajidPas encore d'évaluation
- Bilan Kinesitherapie RespiratorieDocument40 pagesBilan Kinesitherapie Respiratoriecarla garciaPas encore d'évaluation
- 4.2.s2 Cours N 2Document3 pages4.2.s2 Cours N 2mido78100% (1)
- Accidents Du SportDocument20 pagesAccidents Du SportYassine DrhPas encore d'évaluation
- BB08 PDFDocument6 pagesBB08 PDFTamer Mohamed SalehPas encore d'évaluation
- Examen Physique Du Nouveau-Ne ThiaDocument109 pagesExamen Physique Du Nouveau-Ne Thiajeanfrancoisgerline70Pas encore d'évaluation
- Détresses Respiratoire Du Nouveau NéDocument13 pagesDétresses Respiratoire Du Nouveau NésarahPas encore d'évaluation
- I) Evaluation Des Resssources Partie A: Evaluation Des Savoirs Exercice 1: Questions À Choix Multiples (QCM)Document8 pagesI) Evaluation Des Resssources Partie A: Evaluation Des Savoirs Exercice 1: Questions À Choix Multiples (QCM)mahopfranck092Pas encore d'évaluation
- 7 Tissu Musculaire Polycope 2020Document15 pages7 Tissu Musculaire Polycope 2020randomPas encore d'évaluation
- Apo A1Document2 pagesApo A1Souad DahmaPas encore d'évaluation
- Kalenji - Preparer Un 10km en 60 Minutes Environ - 17230 PDFDocument25 pagesKalenji - Preparer Un 10km en 60 Minutes Environ - 17230 PDFYassine BoumehdiPas encore d'évaluation
- Fiches D'anatomie - MBR SupDocument122 pagesFiches D'anatomie - MBR SupAmghidh100% (1)
- S217 Chapitre 6 Les MosissuresDocument64 pagesS217 Chapitre 6 Les MosissuresBadr GourmahPas encore d'évaluation
- Acrosyndromes Vasculaires 2014Document6 pagesAcrosyndromes Vasculaires 2014mitroisergiuPas encore d'évaluation
- L'état de ChocDocument5 pagesL'état de ChocJeager ErenPas encore d'évaluation
- C11. Physiologie Du Pancéas EndocrineDocument8 pagesC11. Physiologie Du Pancéas EndocrineAssarm Albattar KikoPas encore d'évaluation
- EmballageDocument1 pageEmballagedexteralexis1210Pas encore d'évaluation
- Le Livre L'appli Le Livre L'appli Le Livre L'appli Le Livre L'appli Le Livre L'appliDocument34 pagesLe Livre L'appli Le Livre L'appli Le Livre L'appli Le Livre L'appli Le Livre L'appliP100% (2)
- SVT 3ème - L2 - La Digestion Des AlimentsDocument13 pagesSVT 3ème - L2 - La Digestion Des Alimentsjean michael kouakouPas encore d'évaluation
- 6 - Appareil RespiratoireDocument4 pages6 - Appareil RespiratoireClélia RatPas encore d'évaluation
- Étude de La Phytochimie Et Activités Biologique...Document100 pagesÉtude de La Phytochimie Et Activités Biologique...Polo CXPas encore d'évaluation
- Anatomie OculaireDocument26 pagesAnatomie OculaireKhallil OuahlimaPas encore d'évaluation
- Les Sens Et Les Récepteurs Sensoriels Secondaire Alloprof PDFDocument1 pageLes Sens Et Les Récepteurs Sensoriels Secondaire Alloprof PDFLolez BoxoPas encore d'évaluation
- Culture Du Cannabis - Problemes, Carence, ExcesDocument25 pagesCulture Du Cannabis - Problemes, Carence, ExcesXela Zoilrac100% (2)