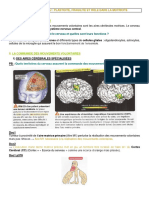
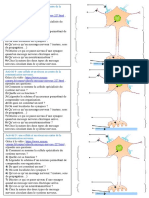
Vous aimerez peut-être aussi
- Neurobiologie: Les Grands Articles d'UniversalisD'EverandNeurobiologie: Les Grands Articles d'UniversalisPas encore d'évaluation
- Cours Et TD CHP II Motricité VolontaireDocument5 pagesCours Et TD CHP II Motricité VolontaireYzyz Ehydsjz100% (1)
- Cours Et TD CHP II Motricité VolontaireDocument5 pagesCours Et TD CHP II Motricité VolontaireYzyz EhydsjzPas encore d'évaluation
- Motricité humaine - Tome 1: Fondements et applications pédagogiquesD'EverandMotricité humaine - Tome 1: Fondements et applications pédagogiquesPas encore d'évaluation
- TD Reflexe MyotatiqueDocument4 pagesTD Reflexe MyotatiquelemiaPas encore d'évaluation
- Chapitre 3A Cours Sur Cerveau Et Mouvement VolontaireDocument3 pagesChapitre 3A Cours Sur Cerveau Et Mouvement Volontairealice.maranberPas encore d'évaluation
- Neuroanatomie Funciones-LexicoDocument69 pagesNeuroanatomie Funciones-LexicoDaniela RiosPas encore d'évaluation
- 003 Eae 404 ESSVTSS7Document3 pages003 Eae 404 ESSVTSS7torkitaherPas encore d'évaluation
- Anatomie FonctionnelleDocument44 pagesAnatomie FonctionnelleAbir SaalaouiPas encore d'évaluation
- 3 Progression Communication NerveuseDocument8 pages3 Progression Communication NerveuseDu charme QuinsPas encore d'évaluation
- Neuroanatomie FonctionnelleDocument14 pagesNeuroanatomie FonctionnelleWilliams SantimePas encore d'évaluation
- Sensorial It ÉDocument5 pagesSensorial It Éjvf4prjq6wPas encore d'évaluation
- Le Muscle Strié Squelettique: Georges CAZORLADocument27 pagesLe Muscle Strié Squelettique: Georges CAZORLAAlphonse AnaniPas encore d'évaluation
- SN06TEWB0522 Retenir-PartieIDocument10 pagesSN06TEWB0522 Retenir-PartieIremandasarahPas encore d'évaluation
- COURS Chapitre 1Document5 pagesCOURS Chapitre 1Aminou MounirouPas encore d'évaluation
- Cours Neurones Synapses PDFDocument82 pagesCours Neurones Synapses PDFSofiane Salem100% (1)
- Electromyographie Et Potentiels Evoqués Dans Les Pathologies de L'Appareil LocomoteurDocument20 pagesElectromyographie Et Potentiels Evoqués Dans Les Pathologies de L'Appareil LocomoteurAicha BenzagoutaPas encore d'évaluation
- Tonus MotriciteDocument15 pagesTonus MotricitecharbelhannaPas encore d'évaluation
- T3B2 Motricite Volonte Et Plasticite CerebraleDocument51 pagesT3B2 Motricite Volonte Et Plasticite CerebralelabsoneducationPas encore d'évaluation
- Cortex Moteur 2019Document49 pagesCortex Moteur 2019Sofiane SalemPas encore d'évaluation
- FetchDocument13 pagesFetchAhmed RekikPas encore d'évaluation
- Tspe Chap2 Le CerveauDocument11 pagesTspe Chap2 Le Cerveauhervy margauxPas encore d'évaluation
- Exercice ReflexeDocument11 pagesExercice ReflexeBathie SarrPas encore d'évaluation
- Cours Système NerveuxDocument25 pagesCours Système NerveuxAblaye100% (1)
- Organisation Generale Du Système NerveuxkDocument21 pagesOrganisation Generale Du Système NerveuxkTETY FREJUS BEUGREPas encore d'évaluation
- UE2 Tome 4 Sur 4Document145 pagesUE2 Tome 4 Sur 4mathieu.lanier.2004Pas encore d'évaluation
- TD Organisation Du Systeme NerveuxDocument8 pagesTD Organisation Du Systeme NerveuxdiarraPas encore d'évaluation
- Le Fonctionnement Du Systeme NerveuxDocument11 pagesLe Fonctionnement Du Systeme NerveuxSadi KoukouPas encore d'évaluation
- L'Acte MoteurDocument10 pagesL'Acte MoteurJulien MunozPas encore d'évaluation
- Chapitre 04 - Réseaux de NeuronesDocument17 pagesChapitre 04 - Réseaux de NeuronesAhmed Ayoub OsmaniPas encore d'évaluation
- Utilisation Des Cartes Auto-Organisatrices de Kohonen Dans La Recherche Documentaire PDFDocument14 pagesUtilisation Des Cartes Auto-Organisatrices de Kohonen Dans La Recherche Documentaire PDFStephane MbiaPas encore d'évaluation
- Résumé PsychophysioDocument34 pagesRésumé PsychophysioJoël OdiPas encore d'évaluation
- Chapitre 9 - Réflexe Myotatique Et Message NerveuxDocument10 pagesChapitre 9 - Réflexe Myotatique Et Message NerveuxYousra INDIAPas encore d'évaluation
- Fiche 8 - SVT - TS2 + CorrigeDocument1 pageFiche 8 - SVT - TS2 + CorrigeSeydina Wade100% (4)
- Chapitre 4 - IEF Neuromusculaire (EMG)Document43 pagesChapitre 4 - IEF Neuromusculaire (EMG)zenazel miloudPas encore d'évaluation
- 123 Acte Moteur Congres Eps 76Document17 pages123 Acte Moteur Congres Eps 76Sadi KoukouPas encore d'évaluation
- Psychophysio PDF LMD 2015Document46 pagesPsychophysio PDF LMD 2015AliBenMessaoudPas encore d'évaluation
- Article Comm 0588-8018 1972 Num 18 1 1257Document12 pagesArticle Comm 0588-8018 1972 Num 18 1 1257lmehdi237Pas encore d'évaluation
- EXERCICE de DissertationDocument1 pageEXERCICE de DissertationkingbachsambPas encore d'évaluation
- Cours TIA - Chapitre 2 PDFDocument11 pagesCours TIA - Chapitre 2 PDFHoussine GuePas encore d'évaluation
- Th2 Chap 3 Act 9Document1 pageTh2 Chap 3 Act 9khayrou 692Pas encore d'évaluation
- Isha Metatron Hunter GR Pro User Manual Training 2023 Francais FrenchDocument100 pagesIsha Metatron Hunter GR Pro User Manual Training 2023 Francais FrenchManuela PravettoniPas encore d'évaluation
- Neurophysiologie PDFDocument140 pagesNeurophysiologie PDFPascal Kasero0% (1)
- 11 - Cortex MoteurDocument46 pages11 - Cortex Moteurkassirmanal24Pas encore d'évaluation
- TP Le Reflexe Myotatique Sujet Et CorrigeDocument3 pagesTP Le Reflexe Myotatique Sujet Et Corrigeusless.fairyPas encore d'évaluation
- Introduction Aux Reseaux de NeuronesDocument31 pagesIntroduction Aux Reseaux de NeuronesIbrahim RanaoPas encore d'évaluation
- Musique (Enregistré Automatiquement)Document49 pagesMusique (Enregistré Automatiquement)Siham AmessaghrouPas encore d'évaluation
- 19a Transmis SynaptDocument15 pages19a Transmis SynaptTalel TouaitiPas encore d'évaluation
- 3e - T3 - C2 - Act 1 - La Commande Du MouvementDocument1 page3e - T3 - C2 - Act 1 - La Commande Du MouvementOrlane JaquinandiPas encore d'évaluation
- 3 EmgDocument12 pages3 EmgIchrak HizemPas encore d'évaluation
- UntitledDocument2 pagesUntitlednawal benyPas encore d'évaluation
- La Communication Nerveuse: Les PrérequisDocument24 pagesLa Communication Nerveuse: Les PrérequisFerssiwi OmarPas encore d'évaluation
- Réseaux de Neurone ArtificielDocument29 pagesRéseaux de Neurone ArtificiellebelulePas encore d'évaluation
- Réflexe TspéDocument8 pagesRéflexe TspéambreletallecPas encore d'évaluation
- Le Système Nerveux: Exercice 1Document3 pagesLe Système Nerveux: Exercice 1NoraMatou100% (6)
- My DocumentsDocument12 pagesMy DocumentsGneneziPas encore d'évaluation
- SVT Le Reflexe Myotatique PDFDocument7 pagesSVT Le Reflexe Myotatique PDFAnonymous TrrvM8Pas encore d'évaluation
- Chapitre 1Document3 pagesChapitre 1leny3011Pas encore d'évaluation
- Nouveau Rapport MLADocument20 pagesNouveau Rapport MLAzicozo012Pas encore d'évaluation
- La Gestion de La Paie PDFDocument96 pagesLa Gestion de La Paie PDFYacine AyatPas encore d'évaluation
- P 20133329Document142 pagesP 20133329andoPas encore d'évaluation
- Cours 2Document4 pagesCours 2free fire xyahiaxPas encore d'évaluation
- Série de TD 03Document2 pagesSérie de TD 03BilaPas encore d'évaluation
- Crise 2000 de La Cote D'ivoirDocument4 pagesCrise 2000 de La Cote D'ivoirballa pierre koivoguiPas encore d'évaluation
- Magir1 Devoir Ipv6 2014 2015Document3 pagesMagir1 Devoir Ipv6 2014 2015assouoPas encore d'évaluation
- Support Stratégies Des Firmes MultinationalesDocument38 pagesSupport Stratégies Des Firmes MultinationalesmariaPas encore d'évaluation
- (Free Scores - Com) - 039 Guessan Gna Houa Jean Claude Sanctus Sainte Bernadette 75050Document2 pages(Free Scores - Com) - 039 Guessan Gna Houa Jean Claude Sanctus Sainte Bernadette 75050Edmond NIREMAPas encore d'évaluation
- Problématique: Comment Marivaux Réussit-Il À Mettre en Abyme Les Différents Niveaux de Stratagèmes Mis en Œuvre Dans Cette Scène ?Document3 pagesProblématique: Comment Marivaux Réussit-Il À Mettre en Abyme Les Différents Niveaux de Stratagèmes Mis en Œuvre Dans Cette Scène ?Louis BoulayPas encore d'évaluation
- Moi GianniDocument1 pageMoi GianniIlham BnPas encore d'évaluation
- Douville O Enfants Des RuesDocument33 pagesDouville O Enfants Des RuesJean Rigobert MbengPas encore d'évaluation
- ComptabiliteDocument503 pagesComptabiliteAhmed JebariPas encore d'évaluation
- AlkhawarichtiDocument2 pagesAlkhawarichtisyll.el-hadji-massePas encore d'évaluation
- Comment Prospecter de Nouveaux ClientsDocument10 pagesComment Prospecter de Nouveaux ClientsAyadi NizarPas encore d'évaluation
- Thème 3 Le Cid Evaluation BilanDocument3 pagesThème 3 Le Cid Evaluation BilanYounes Larhrib100% (1)
- Interview-Bounajah Compo 1asDocument3 pagesInterview-Bounajah Compo 1asMaache Nadjet100% (1)
- Qâsim - Al Firansi PDFDocument6 pagesQâsim - Al Firansi PDFmac51100Pas encore d'évaluation
- 4 Atomes MoléculesDocument3 pages4 Atomes MoléculesSidhoum SidPas encore d'évaluation
- Coaching Personnel IndividuelDocument2 pagesCoaching Personnel Individuelmuller senséPas encore d'évaluation
- Roto 16sept 1ed PDFDocument16 pagesRoto 16sept 1ed PDFLa_RotondePas encore d'évaluation
- M01 Métier Et Formation-GE-TEMIDocument43 pagesM01 Métier Et Formation-GE-TEMIltc100% (1)
- PrepECN Item 90 - Infections Naso-Sinusiennes de L'adulte Et de L'enfant - Fiches de Préparation Aux ECN de MédecineDocument7 pagesPrepECN Item 90 - Infections Naso-Sinusiennes de L'adulte Et de L'enfant - Fiches de Préparation Aux ECN de MédecineAdémonla ROUFAÏPas encore d'évaluation
- WTPW-Simon PierreDocument100 pagesWTPW-Simon Pierrekonan kouadio henri franckPas encore d'évaluation
- Ichimoku Timespan Theory Nombre9Document2 pagesIchimoku Timespan Theory Nombre9san RayPas encore d'évaluation
- French Writing - QuestionsDocument13 pagesFrench Writing - QuestionsSujithPas encore d'évaluation
- MasterDocument1 pageMasterMaoukil TachPas encore d'évaluation
- Cor - GEE1 - 1 La Grammaire en Exercices Cahier 1Document71 pagesCor - GEE1 - 1 La Grammaire en Exercices Cahier 1Adriano YamaokaPas encore d'évaluation
- CH 1 Analyse Financière ESGDocument8 pagesCH 1 Analyse Financière ESGOualid OunaceurPas encore d'évaluation
- Histoire de la psychologie scientifique: De la naissance de la psychologie à la neuropsychologie et aux champs d'application les plus actuelsD'EverandHistoire de la psychologie scientifique: De la naissance de la psychologie à la neuropsychologie et aux champs d'application les plus actuelsPas encore d'évaluation
- Améliorer votre mémoire: Un Guide pour l'augmentation de la puissance du cerveau, utilisant des techniques et méthodesD'EverandAméliorer votre mémoire: Un Guide pour l'augmentation de la puissance du cerveau, utilisant des techniques et méthodesÉvaluation : 5 sur 5 étoiles5/5 (2)
- Harmonisation Energétique des Lieux: Habitat et haut-lieux sacrés 2020D'EverandHarmonisation Energétique des Lieux: Habitat et haut-lieux sacrés 2020Évaluation : 2.5 sur 5 étoiles2.5/5 (3)
- Introduction à la psychologie des émotions: De Darwin aux neurosciences: découvrir les émotions et leur mode de fonctionnementD'EverandIntroduction à la psychologie des émotions: De Darwin aux neurosciences: découvrir les émotions et leur mode de fonctionnementPas encore d'évaluation
- Neuropsychologie: Les bases théoriques et pratiques du domaine d'étude (psychologie pour tous)D'EverandNeuropsychologie: Les bases théoriques et pratiques du domaine d'étude (psychologie pour tous)Pas encore d'évaluation
- La vie des abeilles: Prix Nobel de littératureD'EverandLa vie des abeilles: Prix Nobel de littératureÉvaluation : 4 sur 5 étoiles4/5 (41)
- Cahier de jeux de stimulation cognitive: Sujets Alzheimer, désorientés, démences, amnésiesD'EverandCahier de jeux de stimulation cognitive: Sujets Alzheimer, désorientés, démences, amnésiesPas encore d'évaluation
- L' ESPRIT EST SON PROPRE MÉDECIN: Le pouvoir de guérison de la méditationD'EverandL' ESPRIT EST SON PROPRE MÉDECIN: Le pouvoir de guérison de la méditationPas encore d'évaluation
- Hypnose & affirmations positives pour la richesse, l’abondance et la réussite (21 jours pour un nouveau vous)D'EverandHypnose & affirmations positives pour la richesse, l’abondance et la réussite (21 jours pour un nouveau vous)Évaluation : 5 sur 5 étoiles5/5 (1)
- Géologie de l'Amérique: Les Grands Articles d'UniversalisD'EverandGéologie de l'Amérique: Les Grands Articles d'UniversalisPas encore d'évaluation
- Qu'arrive t'il a la terre arable ?: Le problème de l'érosion du sol.D'EverandQu'arrive t'il a la terre arable ?: Le problème de l'érosion du sol.Évaluation : 5 sur 5 étoiles5/5 (1)
- La psychologie de l'anxiété: La connaître pour comprendre ses mécanismes de fonctionnementD'EverandLa psychologie de l'anxiété: La connaître pour comprendre ses mécanismes de fonctionnementPas encore d'évaluation
- Le TDAH chez l’adulte : Comment reconnaître & faire face au TDAH chez l’adulte en 30 étapes faciles.D'EverandLe TDAH chez l’adulte : Comment reconnaître & faire face au TDAH chez l’adulte en 30 étapes faciles.Pas encore d'évaluation
- Mes tempêtes intérieures: 25 témoignages porteurs d'espoir sur la santé mentaleD'EverandMes tempêtes intérieures: 25 témoignages porteurs d'espoir sur la santé mentalePas encore d'évaluation
- Le CODE DE DIEU: Le secret de notre passé, la promesse de notre avenirD'EverandLe CODE DE DIEU: Le secret de notre passé, la promesse de notre avenirÉvaluation : 4 sur 5 étoiles4/5 (23)
- Le Pouvoir De L'Autodiscipline: L’art de tout accomplir sans limiteD'EverandLe Pouvoir De L'Autodiscipline: L’art de tout accomplir sans limiteÉvaluation : 4 sur 5 étoiles4/5 (16)
- La Conscience Et L'Univers Existent Sans Commencement Ni FinD'EverandLa Conscience Et L'Univers Existent Sans Commencement Ni FinPas encore d'évaluation
- Atlas permanent de l'Union européenne: 5e édition revue et augmentéeD'EverandAtlas permanent de l'Union européenne: 5e édition revue et augmentéePas encore d'évaluation
- La thérapie libre: La Thérapie de LIBération et Résilience EmotionnelleD'EverandLa thérapie libre: La Thérapie de LIBération et Résilience EmotionnelleÉvaluation : 5 sur 5 étoiles5/5 (1)
- Pourquoi suis-je différent?: Accepter et découvrir la puissance de l'hypersensibilitéD'EverandPourquoi suis-je différent?: Accepter et découvrir la puissance de l'hypersensibilitéPas encore d'évaluation
- Connaître ses réactions émotionnelles, pour maîtriser ses émotionsD'EverandConnaître ses réactions émotionnelles, pour maîtriser ses émotionsPas encore d'évaluation
- La motivation: Voyage au cœur des comportements motivés, de l'étude des processus internes aux théories neuropsychologiques les plus récentesD'EverandLa motivation: Voyage au cœur des comportements motivés, de l'étude des processus internes aux théories neuropsychologiques les plus récentesPas encore d'évaluation