Vous aimerez peut-être aussi
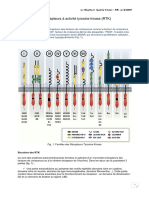
- Les Récepteurs À Activité Tyrosine KinaseDocument10 pagesLes Récepteurs À Activité Tyrosine KinaseFatima Zahra100% (1)
- Membranes cellulaires: Les Grands Articles d'UniversalisD'EverandMembranes cellulaires: Les Grands Articles d'UniversalisPas encore d'évaluation
- Récepteurs À Activité Tyrosine Kinase DefDocument12 pagesRécepteurs À Activité Tyrosine Kinase DefFatima ZahraPas encore d'évaluation
- Recepteur de L'insuline G1Document12 pagesRecepteur de L'insuline G1페 리알Pas encore d'évaluation
- Chap II Récepteurs Membranaires Et Molécules de Signalisation IntracellulaireDocument28 pagesChap II Récepteurs Membranaires Et Molécules de Signalisation IntracellulaireAbdoulaye sebyPas encore d'évaluation
- Cours RéglationDocument33 pagesCours RéglationBADzPas encore d'évaluation
- Recepteurs Associes A Une Tyrosine KinaseDocument13 pagesRecepteurs Associes A Une Tyrosine KinaseFatima ZahraPas encore d'évaluation
- 3eme Biochimie 2019-2020 Mahdab N L3 Bioch COURS DE BIO MOLDocument22 pages3eme Biochimie 2019-2020 Mahdab N L3 Bioch COURS DE BIO MOLry yukPas encore d'évaluation
- Tpe 447 FiniDocument15 pagesTpe 447 FinijndjiendeuPas encore d'évaluation
- Httpselearning Centre-Univ-Mila Dza2024pluginfile Php89198mod - resourcecontent1CHAPITRE20V PDFDocument20 pagesHttpselearning Centre-Univ-Mila Dza2024pluginfile Php89198mod - resourcecontent1CHAPITRE20V PDFariel.2mariemPas encore d'évaluation
- UE2-Intégration - 13 - Communication-IntercellulaireDocument17 pagesUE2-Intégration - 13 - Communication-Intercellulairesun-nee-chan9Pas encore d'évaluation
- Recepteur A Activite Serine Threonine KinaseDocument11 pagesRecepteur A Activite Serine Threonine KinaseFatima ZahraPas encore d'évaluation
- UE2 - Biologie Cellulaire - Le CytosolDocument11 pagesUE2 - Biologie Cellulaire - Le Cytosolsun-nee-chan9Pas encore d'évaluation
- Recepteurs Protéines - Partie 3.2Document49 pagesRecepteurs Protéines - Partie 3.2Anonymous MKSfyYyODPPas encore d'évaluation
- Biotech EnzymatiqueDocument79 pagesBiotech Enzymatiquelahouam nihed100% (1)
- 2.-Biosynthèse Des ProtéinesDocument65 pages2.-Biosynthèse Des Protéinesmido benPas encore d'évaluation
- Cours BMDocument22 pagesCours BMNidhaloo NidPas encore d'évaluation
- Biologie Moleculaire 2Document10 pagesBiologie Moleculaire 2Xzistence100% (3)
- 2023-3BB - Classes RCDocument36 pages2023-3BB - Classes RCIsabella AlmeidaPas encore d'évaluation
- Chap 3Document20 pagesChap 3Radjaa AddPas encore d'évaluation
- Biologie Moléculaire Chapitre 3 REGULATION M1 1Document8 pagesBiologie Moléculaire Chapitre 3 REGULATION M1 1اميرة في عرش ابيPas encore d'évaluation
- 11 Traduction Chap IV Expression Inf GenDocument5 pages11 Traduction Chap IV Expression Inf GenRock In BandPas encore d'évaluation
- Interaction Ligand RecepteurDocument3 pagesInteraction Ligand RecepteurAmina MnPas encore d'évaluation
- Qe BiocellDocument9 pagesQe BiocellASSIA EL MOUSTAID100% (1)
- Ladressage Des Protéines - PDF Dans RERDocument48 pagesLadressage Des Protéines - PDF Dans RERamal ELYOUBIPas encore d'évaluation
- Anti Bio TiqueDocument23 pagesAnti Bio Tiquemina ycPas encore d'évaluation
- Chapitre 5signalisationDocument7 pagesChapitre 5signalisationFatïma ZohraPas encore d'évaluation
- Compartiments Intracell Et Tri Des ProtéinesDocument43 pagesCompartiments Intracell Et Tri Des Protéinesa29921Pas encore d'évaluation
- RibosomeDocument30 pagesRibosomeFatima BENHAFSSAPas encore d'évaluation
- Récept-Enzymes-Cours M1-2023-PCMDocument32 pagesRécept-Enzymes-Cours M1-2023-PCMAggoun FadouaPas encore d'évaluation
- 01 C Interaction Ligand RécepteurDocument39 pages01 C Interaction Ligand RécepteurKhelifi SarraPas encore d'évaluation
- Cytoplasme 4Document27 pagesCytoplasme 4Oumarou KontaPas encore d'évaluation
- Bio Mol L3 MicrobiologieDocument18 pagesBio Mol L3 MicrobiologieimanePas encore d'évaluation
- Biologie Moléculaire Chapitre 1 T Procaryote M1Document8 pagesBiologie Moléculaire Chapitre 1 T Procaryote M1اميرة في عرش ابيPas encore d'évaluation
- Mi To Chon DriesDocument9 pagesMi To Chon Driesamira14.mira83Pas encore d'évaluation
- ExposéDocument23 pagesExposéTheSoul OfWind100% (1)
- Med Genetique-Transcription EucaryotesDocument20 pagesMed Genetique-Transcription EucaryotesRacha MalakPas encore d'évaluation
- TD Signal 2023Document10 pagesTD Signal 2023salym.mourid84Pas encore d'évaluation
- 11 - Chap11-Les Différents Modes D'action Des HormonesDocument6 pages11 - Chap11-Les Différents Modes D'action Des HormonesYassine HassanePas encore d'évaluation
- Biochimie Des Protéines ObjectifsDocument14 pagesBiochimie Des Protéines ObjectifsYwenn SAMYNPas encore d'évaluation
- Correction Devoir de Génétique MoléculaireDocument3 pagesCorrection Devoir de Génétique MoléculairessggnfPas encore d'évaluation
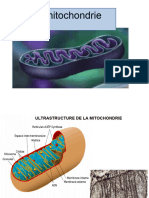
- Cyto1an Mitochondrie2022aouatiDocument12 pagesCyto1an Mitochondrie2022aouatiAy ManPas encore d'évaluation
- 1 TranscriptionDocument12 pages1 TranscriptionYounes YouyouPas encore d'évaluation
- Récepteurs MembranairesDocument56 pagesRécepteurs MembranairesAnonymous MKSfyYyODPPas encore d'évaluation
- ULB EXAMEN BiocellDocument6 pagesULB EXAMEN Biocellkiki02100% (1)
- 01 - La Communication IntercellulaireDocument8 pages01 - La Communication IntercellulaireHadil BenslamaPas encore d'évaluation
- Chap - 5 - Le CytosqueletteDocument12 pagesChap - 5 - Le CytosqueletteMoeanau GrangerPas encore d'évaluation
- La Liaison Aux RécepteursDocument8 pagesLa Liaison Aux RécepteursIsmaelPas encore d'évaluation
- 3 Synthèse Protéique-2Document12 pages3 Synthèse Protéique-2Sagacious IvejutenPas encore d'évaluation
- Cibles Moléculaires Et Élaboration de La Réponse 2024Document3 pagesCibles Moléculaires Et Élaboration de La Réponse 2024kaouaneamdjadPas encore d'évaluation
- Httpsstaff - Univ-Batna2.dzsitesdefaultfilesabdessemed Samirafileschapitre 11 PDFDocument12 pagesHttpsstaff - Univ-Batna2.dzsitesdefaultfilesabdessemed Samirafileschapitre 11 PDFariel.2mariemPas encore d'évaluation
- Null PDFDocument26 pagesNull PDFAziz OualidPas encore d'évaluation
- Transcription 21 - 11 - 2023Document55 pagesTranscription 21 - 11 - 2023Elmahdi BOUHFIDPas encore d'évaluation
- Les ARNm Cytoplasmiques1Document9 pagesLes ARNm Cytoplasmiques1Sokhna gurlPas encore d'évaluation
- Partie I. CH1.3. Altérations Et Mécanismes de Réparation Du Génome Bactérien ETUDDocument21 pagesPartie I. CH1.3. Altérations Et Mécanismes de Réparation Du Génome Bactérien ETUDAziza BoulberhanePas encore d'évaluation
- Chap 7Document22 pagesChap 7Ayoub LilouPas encore d'évaluation
- Physio1an-Interactions Ligand Recepteur2021bougrida BoussoufDocument24 pagesPhysio1an-Interactions Ligand Recepteur2021bougrida BoussoufGI 2 Session 2018-2019Pas encore d'évaluation
- Chapitre 3Document5 pagesChapitre 3bouchakour meryemPas encore d'évaluation
- Interactions Ligand - Recepteur 2023-2024Document29 pagesInteractions Ligand - Recepteur 2023-2024khaldiniheddoc23Pas encore d'évaluation
- Denutrition DrlimontaDocument18 pagesDenutrition DrlimontaAbdoulaye sebyPas encore d'évaluation
- Exploration Du Metabolisme GlucidiqueDocument40 pagesExploration Du Metabolisme GlucidiqueAbdoulaye sebyPas encore d'évaluation
- Chapitre IV Les Etudes Transversales P Duhaut J SchmidtDocument16 pagesChapitre IV Les Etudes Transversales P Duhaut J SchmidtAbdoulaye sebyPas encore d'évaluation
- Exploration Du Métabolisme HépatiqueDocument40 pagesExploration Du Métabolisme HépatiqueAbdoulaye sebyPas encore d'évaluation
- Exploration de Léquilibre HydrominéralDocument37 pagesExploration de Léquilibre HydrominéralAbdoulaye sebyPas encore d'évaluation
- Centrifugation Et Ultracentrifugation-ConvertiDocument10 pagesCentrifugation Et Ultracentrifugation-ConvertiAbdoulaye sebyPas encore d'évaluation
- Contrôle de Qualité Microbiologique CoursDocument12 pagesContrôle de Qualité Microbiologique CoursAbdoulaye seby100% (1)
- Biochimie 2eme Emed 2016-2017Document7 pagesBiochimie 2eme Emed 2016-2017Louiza MecheloufPas encore d'évaluation
- Infographie Carrefour - OGM, Ce Qu Il Faut Savoir - Page 1 - Mars 2004 PDFDocument1 pageInfographie Carrefour - OGM, Ce Qu Il Faut Savoir - Page 1 - Mars 2004 PDFJustinePas encore d'évaluation
- Genesis 1Document6 pagesGenesis 1Neydimar HernandezPas encore d'évaluation
- SE Cours 3OSDocument22 pagesSE Cours 3OSLara FarquharsonPas encore d'évaluation
- Calalogue 2018 Converti PDFDocument149 pagesCalalogue 2018 Converti PDFLili BioPas encore d'évaluation
- 1469 - 1S - TP14 Contrôle Par AuxineDocument1 page1469 - 1S - TP14 Contrôle Par Auxineelhidhab2014Pas encore d'évaluation
- Diapo - Gamétog & Fécond DéfinitifDocument14 pagesDiapo - Gamétog & Fécond DéfinitifL kPas encore d'évaluation
- tp7 CorrigeDocument4 pagestp7 CorrigeEmmanuel KoffiPas encore d'évaluation
- COLLEGE MARIAMA DEVOIR Jan23 TDDocument2 pagesCOLLEGE MARIAMA DEVOIR Jan23 TDMaria Alielhadjiabdou100% (1)
- Cours Palléontologie GS3 2022 2023.ppt Pour ÉtudiantsDocument101 pagesCours Palléontologie GS3 2022 2023.ppt Pour ÉtudiantsMed LeminPas encore d'évaluation
- Formation Biologie MoléculaireDocument42 pagesFormation Biologie MoléculaireRym BldjPas encore d'évaluation
- Développement Des Leçons de La Série: Leçon 1 Le Vivant Et Le Non-VivantDocument38 pagesDéveloppement Des Leçons de La Série: Leçon 1 Le Vivant Et Le Non-VivantMoh'dTfailiPas encore d'évaluation
- Catalogue de La Flore Vasculaire de La Région Rhône-Alpes PDFDocument196 pagesCatalogue de La Flore Vasculaire de La Région Rhône-Alpes PDFDavidPas encore d'évaluation
- Guérir Alzheimer - Comprendre Et Agir À Temps (Questions de Santé) (French EDocument284 pagesGuérir Alzheimer - Comprendre Et Agir À Temps (Questions de Santé) (French Ejbaltagi100% (2)
- 25 CetogeneseDocument5 pages25 CetogeneseSerigne Sohibou GayePas encore d'évaluation
- ExposéDocument3 pagesExposéIkramPas encore d'évaluation
- Techniques de Coloration Pour Etudier La Cellule (2019)Document2 pagesTechniques de Coloration Pour Etudier La Cellule (2019)almnaouarPas encore d'évaluation
- L Entraide L Autre Loi de La Jungle - Pa PDFDocument214 pagesL Entraide L Autre Loi de La Jungle - Pa PDFWen Rolland100% (1)
- Cours Etudiants V2 2016Document44 pagesCours Etudiants V2 2016Zaidou RoambaPas encore d'évaluation
- Cours10 La Phyto-Remédiation Du SolDocument5 pagesCours10 La Phyto-Remédiation Du SolMer IemPas encore d'évaluation
- Bio P1 TZ1Document25 pagesBio P1 TZ1Martina GuisadoPas encore d'évaluation
- Les Feuilles Des Monocotylédones Et Des DicotylédonesDocument2 pagesLes Feuilles Des Monocotylédones Et Des DicotylédonesMoussaba DoumbouyaPas encore d'évaluation
- S Out EnanceDocument59 pagesS Out EnanceRose RosePas encore d'évaluation
- Programme Des Cours Résidanat FAC ALGERDocument49 pagesProgramme Des Cours Résidanat FAC ALGERBilel BouzeredPas encore d'évaluation
- Biochimie-MétaboliqueDocument18 pagesBiochimie-MétaboliqueLhoussaine MaskourPas encore d'évaluation
- Génie Genet. Exercices - 22-23Document36 pagesGénie Genet. Exercices - 22-23Hanae Benhaida100% (1)
- Améliorer votre mémoire: Un Guide pour l'augmentation de la puissance du cerveau, utilisant des techniques et méthodesD'EverandAméliorer votre mémoire: Un Guide pour l'augmentation de la puissance du cerveau, utilisant des techniques et méthodesÉvaluation : 5 sur 5 étoiles5/5 (2)
- Cahier de jeux de stimulation cognitive: Sujets Alzheimer, désorientés, démences, amnésiesD'EverandCahier de jeux de stimulation cognitive: Sujets Alzheimer, désorientés, démences, amnésiesPas encore d'évaluation
- Les 10 Secrets pour une Vie Plus Heureuse avec la Maladie de ParkinsonD'EverandLes 10 Secrets pour une Vie Plus Heureuse avec la Maladie de ParkinsonPas encore d'évaluation
- Manuel pour les débutants Fabriquez des savons naturelsD'EverandManuel pour les débutants Fabriquez des savons naturelsÉvaluation : 3 sur 5 étoiles3/5 (2)
- La thérapie libre: La Thérapie de LIBération et Résilience EmotionnelleD'EverandLa thérapie libre: La Thérapie de LIBération et Résilience EmotionnelleÉvaluation : 5 sur 5 étoiles5/5 (1)
- Histoire de la psychologie scientifique: De la naissance de la psychologie à la neuropsychologie et aux champs d'application les plus actuelsD'EverandHistoire de la psychologie scientifique: De la naissance de la psychologie à la neuropsychologie et aux champs d'application les plus actuelsPas encore d'évaluation
- La vie des abeilles: Prix Nobel de littératureD'EverandLa vie des abeilles: Prix Nobel de littératureÉvaluation : 4 sur 5 étoiles4/5 (41)
- Nutrition Sportive Fondamentaux et guide pratique du succèsD'EverandNutrition Sportive Fondamentaux et guide pratique du succèsÉvaluation : 5 sur 5 étoiles5/5 (1)
- Guide De Démarrage De L'Apiculture: Le Guide Complet Pour Élever Des AbeillesD'EverandGuide De Démarrage De L'Apiculture: Le Guide Complet Pour Élever Des AbeillesPas encore d'évaluation
- La Conscience Et L'Univers Existent Sans Commencement Ni FinD'EverandLa Conscience Et L'Univers Existent Sans Commencement Ni FinPas encore d'évaluation
- Harmonisation Energétique des Lieux: Habitat et haut-lieux sacrés 2020D'EverandHarmonisation Energétique des Lieux: Habitat et haut-lieux sacrés 2020Évaluation : 2.5 sur 5 étoiles2.5/5 (3)
- Géologie de l'Amérique: Les Grands Articles d'UniversalisD'EverandGéologie de l'Amérique: Les Grands Articles d'UniversalisPas encore d'évaluation
- Microbiologie médicale I: agents pathogènes et microbiome humainD'EverandMicrobiologie médicale I: agents pathogènes et microbiome humainÉvaluation : 5 sur 5 étoiles5/5 (1)
- Pourquoi suis-je différent?: Accepter et découvrir la puissance de l'hypersensibilitéD'EverandPourquoi suis-je différent?: Accepter et découvrir la puissance de l'hypersensibilitéPas encore d'évaluation
- Qu'arrive t'il a la terre arable ?: Le problème de l'érosion du sol.D'EverandQu'arrive t'il a la terre arable ?: Le problème de l'érosion du sol.Évaluation : 5 sur 5 étoiles5/5 (1)
- Le CODE DE DIEU: Le secret de notre passé, la promesse de notre avenirD'EverandLe CODE DE DIEU: Le secret de notre passé, la promesse de notre avenirÉvaluation : 4 sur 5 étoiles4/5 (23)
- Les Graines d'Abricot - Remède de Cancer avec de la Vitamine B17 ?: La Médecine Antique Que l'Industrie Pharmaceutique Moderne CacheD'EverandLes Graines d'Abricot - Remède de Cancer avec de la Vitamine B17 ?: La Médecine Antique Que l'Industrie Pharmaceutique Moderne CachePas encore d'évaluation
- Aux pays d'azur, Nice, Monaco et Menton: Descriptions, histoire, moeurs, légendes, excursions et promenades, flore et faune, itinéraires, renseignements générauxD'EverandAux pays d'azur, Nice, Monaco et Menton: Descriptions, histoire, moeurs, légendes, excursions et promenades, flore et faune, itinéraires, renseignements générauxPas encore d'évaluation
- Transition énergétique ET Changement climatique: Développements et Perspectives d'AvenirD'EverandTransition énergétique ET Changement climatique: Développements et Perspectives d'AvenirPas encore d'évaluation
- Encyclopédie de la magie des cristaux, des pierres précieuses et des métauxD'EverandEncyclopédie de la magie des cristaux, des pierres précieuses et des métauxÉvaluation : 4 sur 5 étoiles4/5 (116)