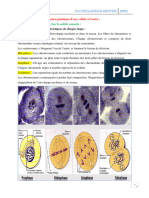
Vous aimerez peut-être aussi
- Cours No 04-Le NoyauDocument15 pagesCours No 04-Le NoyauFeriel Feriel100% (1)
- Chapitre 1 EleveDocument4 pagesChapitre 1 EleveMathis LefrancPas encore d'évaluation
- Chapitre 1 EleveDocument4 pagesChapitre 1 EleveMathis LefrancPas encore d'évaluation
- Blog 2Document1 pageBlog 2tozman1234gta5Pas encore d'évaluation
- Cycle CellulaireDocument37 pagesCycle CellulairegigiPas encore d'évaluation
- Chapitre 2-Le Cycle CellulaireDocument35 pagesChapitre 2-Le Cycle CellulairealmnaouarPas encore d'évaluation
- Cours IV Réplication d'ADN Et Cycle Cellulaire 221115 202921Document6 pagesCours IV Réplication d'ADN Et Cycle Cellulaire 221115 202921Nouha HadjPas encore d'évaluation
- SVT - Origine Du GénotypeDocument10 pagesSVT - Origine Du GénotypefontainePas encore d'évaluation
- Unité 2 RresDocument11 pagesUnité 2 Rresfdell azerPas encore d'évaluation
- SVT - Chapitre 1 Premiere SpecialitéDocument7 pagesSVT - Chapitre 1 Premiere SpecialitéClement LaplacePas encore d'évaluation
- La MitoseDocument4 pagesLa MitoseFDGPas encore d'évaluation
- Chapitre 1 SVT PremiereDocument10 pagesChapitre 1 SVT PremiereInes JaniPas encore d'évaluation
- Chapitre1 Cycle CellulaireDocument17 pagesChapitre1 Cycle CellulaireEnnassiki Med EnnassikiPas encore d'évaluation
- La Nature de L'information GenetiqueDocument6 pagesLa Nature de L'information Genetiqueyassirfaddali2008Pas encore d'évaluation
- Les Mitoses Et MéiosesDocument6 pagesLes Mitoses Et MéiosesTiaret TiaretPas encore d'évaluation
- S.V.T - MifmDocument25 pagesS.V.T - MifmsamPas encore d'évaluation
- Reproduction cellulaire 2023Document23 pagesReproduction cellulaire 2023Jonathan Sola FuentesPas encore d'évaluation
- MITOSE MEIOSE InfoDocument7 pagesMITOSE MEIOSE InfoAHMED DiopPas encore d'évaluation
- Partie 1 Chap 1 Spé SVT Division CellulaireDocument8 pagesPartie 1 Chap 1 Spé SVT Division CellulaireNagwa Elimam100% (2)
- Genetique Cours 2012Document6 pagesGenetique Cours 2012shadowreda18Pas encore d'évaluation
- CoursTH1A Chap1808856045Document10 pagesCoursTH1A Chap1808856045lucariosmandastyleuePas encore d'évaluation
- Chapitre 1-Les Divisions Cellulaires IMPORTANTDocument2 pagesChapitre 1-Les Divisions Cellulaires IMPORTANTphilippePas encore d'évaluation
- Al7sn12tepa0111 Sequence 03Document46 pagesAl7sn12tepa0111 Sequence 03ogmios_940% (1)
- Définition Inf Génétique Et Mendel (PR CHOUNAII)Document5 pagesDéfinition Inf Génétique Et Mendel (PR CHOUNAII)Souad Skali Senhaji100% (1)
- CytogénétiqueDocument33 pagesCytogénétiqueFaiza HPas encore d'évaluation
- SVT Chapitre 1Document4 pagesSVT Chapitre 1scoccimarro.sarahPas encore d'évaluation
- Le Cycle CellulaireDocument2 pagesLe Cycle CellulaireJoëlly CesairePas encore d'évaluation
- MITOSEDocument6 pagesMITOSELarbi SalihPas encore d'évaluation
- Activités Génétique 1 BACSMFDocument24 pagesActivités Génétique 1 BACSMFHamza AmjahdiPas encore d'évaluation
- (Suite) GENETIQUE 1BACSMDocument11 pages(Suite) GENETIQUE 1BACSMnnadanada2901Pas encore d'évaluation
- Cours Etudiants V2 2016Document44 pagesCours Etudiants V2 2016Zaidou RoambaPas encore d'évaluation
- 02 - Gamétogenèse GDocument50 pages02 - Gamétogenèse GIslęm OuaribPas encore d'évaluation
- TD N1 Le Cycle CellulaireDocument6 pagesTD N1 Le Cycle Cellulairemaysoun100% (1)
- TD de GénétiqueDocument22 pagesTD de GénétiqueHakim Rahmani100% (1)
- EMBRYOLOGIE NOTES Final PDFDocument32 pagesEMBRYOLOGIE NOTES Final PDFmadaulePas encore d'évaluation
- Cycle Et Divisions - 2022Document43 pagesCycle Et Divisions - 2022youssouf traorePas encore d'évaluation
- Le Cycle Cellulaire PDFDocument1 pageLe Cycle Cellulaire PDFcapou34Pas encore d'évaluation
- Rappels Indispensables de GénétiqueDocument10 pagesRappels Indispensables de Génétiquebakiclio5Pas encore d'évaluation
- Classeur Guide at 2PC Biof - Info Génétique S1Document13 pagesClasseur Guide at 2PC Biof - Info Génétique S1micproteoPas encore d'évaluation
- Chapitre 6 La MitoseDocument5 pagesChapitre 6 La MitoseMeghane TagliaroPas encore d'évaluation
- BDR - FICHE - Cours 1 Mitose Et MéioseDocument15 pagesBDR - FICHE - Cours 1 Mitose Et Méiosesurgeon forgeronPas encore d'évaluation
- Capture D'écran . 2023-11-20 À 08.21.59Document6 pagesCapture D'écran . 2023-11-20 À 08.21.59bouidirbadisPas encore d'évaluation
- Cours 3 Le Cycle Cellulaire PDFDocument17 pagesCours 3 Le Cycle Cellulaire PDFmalak kerakbiPas encore d'évaluation
- Structure Du Noyau. La Division CellulaireDocument6 pagesStructure Du Noyau. La Division CellulaireAziz ChaiebPas encore d'évaluation
- Biologie Cellulaire IIIDocument12 pagesBiologie Cellulaire IIIInas HassanatPas encore d'évaluation
- Chap2 - Cours Physiologie Cellulaire Ufr Stapv - Ussein 2Document60 pagesChap2 - Cours Physiologie Cellulaire Ufr Stapv - Ussein 2sowcapo2004Pas encore d'évaluation
- Bilan Chapitre 2 La Replication de l' Adn Et Variabilite GenetiqueDocument3 pagesBilan Chapitre 2 La Replication de l' Adn Et Variabilite GenetiqueAymane JiraouiPas encore d'évaluation
- La Division Cellulaire Chez Les Eucaryotes Cours de 1ere - SVTDocument1 pageLa Division Cellulaire Chez Les Eucaryotes Cours de 1ere - SVTkuilokrPas encore d'évaluation
- 2éme Partie BiologieDocument31 pages2éme Partie Biologiecamillemwali943Pas encore d'évaluation
- SVT 2nd C - L10 - Lévolution de Léquipement Chromosomique Dune CelluleDocument20 pagesSVT 2nd C - L10 - Lévolution de Léquipement Chromosomique Dune Cellulefranckkouadio2005Pas encore d'évaluation
- Acides Nucléiques Et Synthése de ProtéineDocument112 pagesAcides Nucléiques Et Synthése de ProtéineSarra MoussaouiPas encore d'évaluation
- CHAPITRE II CYCLES DE VIEDocument5 pagesCHAPITRE II CYCLES DE VIEevident 2014Pas encore d'évaluation
- Notion de Linfo Géné Et Sonex SHDocument14 pagesNotion de Linfo Géné Et Sonex SHhajar hajoraPas encore d'évaluation
- COURS INFORMATION GENETIQUE - SuiteDocument9 pagesCOURS INFORMATION GENETIQUE - SuiteMohamed MoustakimPas encore d'évaluation
- Cours Etudiants pdf160215 PDFDocument38 pagesCours Etudiants pdf160215 PDFHichem HammadiPas encore d'évaluation
- GENETIQUE Chapitre 2Document114 pagesGENETIQUE Chapitre 2LectorPas encore d'évaluation
- Chapitre II - Le Cycle CellulaireDocument6 pagesChapitre II - Le Cycle CellulaireMouna DocPas encore d'évaluation
- UNITÉ 2 Corrigé PDFDocument10 pagesUNITÉ 2 Corrigé PDFIrene PGPas encore d'évaluation
- Le Cycle CellulaireDocument16 pagesLe Cycle CellulaireyoussefPas encore d'évaluation
- Cycle Cellulaire RESUMEDocument27 pagesCycle Cellulaire RESUMEmounia.34Pas encore d'évaluation
- 5-Cycle ÇDocument25 pages5-Cycle ÇMohamed OuHouissPas encore d'évaluation
- SVT - 3BDocument68 pagesSVT - 3BsolalslkkPas encore d'évaluation
- Etincelle 2bac SC-PC SVT Sem1Document39 pagesEtincelle 2bac SC-PC SVT Sem1crmefgroupesvt2Pas encore d'évaluation
- 2éme Partie BiologieDocument31 pages2éme Partie Biologiecamillemwali943Pas encore d'évaluation
- 3 - Division CellulaireDocument5 pages3 - Division CellulaireWi SsalPas encore d'évaluation
- Module de Biologie Animale CoursDocument22 pagesModule de Biologie Animale CoursMerouane GhenamPas encore d'évaluation
- SVT 2nd C - L9 - La Division CellulaireDocument13 pagesSVT 2nd C - L9 - La Division CellulaireTouré Kiyofo HabibPas encore d'évaluation
- Cours N°1 - Division Cellulaire Et Rôle Du CytosqueletteDocument16 pagesCours N°1 - Division Cellulaire Et Rôle Du Cytosquelettedyhp2pswsrPas encore d'évaluation
- Cycle Partie2 1diaDocument32 pagesCycle Partie2 1diaAliBenAhmedPas encore d'évaluation
- 2020, PCF, Inf Génétique, ProfDocument97 pages2020, PCF, Inf Génétique, Profasmae1aghbalouPas encore d'évaluation
- Copie2 TWODocument7 pagesCopie2 TWOKhalid Shorts0% (1)
- La Division Celllulaire Des EucaryotesDocument8 pagesLa Division Celllulaire Des EucaryoteselassadPas encore d'évaluation
- Biologie Cellulaire IIIDocument12 pagesBiologie Cellulaire IIIInas HassanatPas encore d'évaluation
- MitoseDocument10 pagesMitoseOumarou KontaPas encore d'évaluation
- Support - Cours de Biologie - Niveau I - TC - ISAGO - Décembre 2022Document74 pagesSupport - Cours de Biologie - Niveau I - TC - ISAGO - Décembre 2022philatine04Pas encore d'évaluation
- L9 Division CellulaireDocument15 pagesL9 Division CellulaireSamuelPas encore d'évaluation
- BDR - FICHE - Cours 1 Mitose Et MéioseDocument15 pagesBDR - FICHE - Cours 1 Mitose Et Méiosesurgeon forgeronPas encore d'évaluation
- Le Cytosquelette RésuméDocument5 pagesLe Cytosquelette RésuméEva BrovedaniPas encore d'évaluation
- Fichier Produit 2219Document9 pagesFichier Produit 2219BOULAMA HASSANA MASSAWPas encore d'évaluation
- Chapitres 1-2 2024Document140 pagesChapitres 1-2 2024CHAIMAE AFIFPas encore d'évaluation
- Cycle CellulaireDocument29 pagesCycle CellulairetinaPas encore d'évaluation
- Division CellulaireDocument2 pagesDivision CellulaireMaya MayaPas encore d'évaluation
- FICHE 1 Cycle CellulaireDocument6 pagesFICHE 1 Cycle CellulaireAnissa BoumaaroufPas encore d'évaluation
- Correction - TD2 - BI - 125 - Structure - Et Fonction - de La - Cellule - AnimaleDocument5 pagesCorrection - TD2 - BI - 125 - Structure - Et Fonction - de La - Cellule - AnimaleBerthe Aristide Ngue NkongoPas encore d'évaluation
- Division CellulaireDocument1 pageDivision CellulaireDi BenamoPas encore d'évaluation
- Définitions 2 - Nature de L'information Génétique.Document2 pagesDéfinitions 2 - Nature de L'information Génétique.Anime- fansPas encore d'évaluation