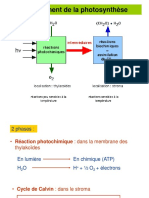
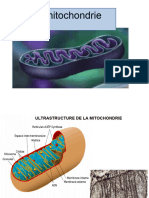
Vous aimerez peut-être aussi
- PhotosynthèseDocument19 pagesPhotosynthèseProost N'guessanPas encore d'évaluation
- La Photosynthèse PDFDocument10 pagesLa Photosynthèse PDFstingx50% (2)
- La Photosynthèse Est Le Processus Responsable de La Transformation de LDocument6 pagesLa Photosynthèse Est Le Processus Responsable de La Transformation de Lamy thiaoPas encore d'évaluation
- Cours-Photosynthse 2022Document68 pagesCours-Photosynthse 2022manalchannel5Pas encore d'évaluation
- Chapitre3 Physiologie VégétaleDocument22 pagesChapitre3 Physiologie Végétaleg69wwpfw5kPas encore d'évaluation
- PhotosyntheseDocument7 pagesPhotosynthesesaraPas encore d'évaluation
- La PhotosyntheseDocument24 pagesLa Photosynthesetoto TOTOROTOPas encore d'évaluation
- Physiologie Vegetale Cours 5photosynthese2Document12 pagesPhysiologie Vegetale Cours 5photosynthese2SALAH-EDDINE AZIZIPas encore d'évaluation
- Photos Yn TheseDocument40 pagesPhotos Yn TheseLINDOR KOUMEPas encore d'évaluation
- COURS BC-6 Chloroplaste - 230805 - 082114Document34 pagesCOURS BC-6 Chloroplaste - 230805 - 082114Mlle BushaPas encore d'évaluation
- Physiologie Vegetale 2 S4Document20 pagesPhysiologie Vegetale 2 S4hassan aitPas encore d'évaluation
- Photosynthèse ELBHAR IBRAHIMDocument60 pagesPhotosynthèse ELBHAR IBRAHIMzah ahmPas encore d'évaluation
- La Photosynthese Cours 1Document7 pagesLa Photosynthese Cours 1essatijihanePas encore d'évaluation
- PhotosynthèseDocument22 pagesPhotosynthèseAyoub Lakhdari0% (1)
- Photosynthèse: Les Grands Articles d'UniversalisD'EverandPhotosynthèse: Les Grands Articles d'UniversalisPas encore d'évaluation
- Energie, Anabolisme, CatabolismeDocument5 pagesEnergie, Anabolisme, CatabolismeMohemed MohamedPas encore d'évaluation
- Quelles Sont Les 2 Phases de La PhotosynthèseDocument7 pagesQuelles Sont Les 2 Phases de La PhotosynthèseMasseylasse EvaPas encore d'évaluation
- Photosynthese 2 S3 Licence HorticultureDocument43 pagesPhotosynthese 2 S3 Licence HorticultureHicham Ait BouhouPas encore d'évaluation
- LBIO1111 CH Part2 2023Document30 pagesLBIO1111 CH Part2 2023jean.bulinckx05Pas encore d'évaluation
- Energie, Anabolisme D'énergie (Photosythèse)Document5 pagesEnergie, Anabolisme D'énergie (Photosythèse)Chawki MokademPas encore d'évaluation
- Photosynthese PascalDocument55 pagesPhotosynthese Pascalmeno donPas encore d'évaluation
- Bio Cell (Semaine 4)Document77 pagesBio Cell (Semaine 4)Maryem BouzebbarPas encore d'évaluation
- La PhotosynthèseDocument35 pagesLa PhotosynthèseELEULJ karimPas encore d'évaluation
- Matière Microorganismes PhotosynthétiquesDocument114 pagesMatière Microorganismes Photosynthétiqueshayet GhalemPas encore d'évaluation
- 3.1 Rappels Sur Les Types Trophiques: 3) La PhotosynthèseDocument23 pages3.1 Rappels Sur Les Types Trophiques: 3) La PhotosynthèseAmanda LaihongtingPas encore d'évaluation
- PhotosyntheseDocument40 pagesPhotosynthesestingx50% (2)
- Université Abdelhamid Ibn Badis-Mostaganem Faculté Des Sciences Exactes Et de L'informatique Département de PhysiqueDocument13 pagesUniversité Abdelhamid Ibn Badis-Mostaganem Faculté Des Sciences Exactes Et de L'informatique Département de PhysiqueLina LA BlondePas encore d'évaluation
- Diversité Métabolique Des MicroorganismesDocument4 pagesDiversité Métabolique Des MicroorganismesAzalais Garcia CrosPas encore d'évaluation
- Document Sans TitreDocument2 pagesDocument Sans Titrelovely glowyPas encore d'évaluation
- Tp3 Phase PhotochimiqueDocument2 pagesTp3 Phase PhotochimiquejamjamalPas encore d'évaluation
- Mecanisme de La Photosynthese CoursDocument15 pagesMecanisme de La Photosynthese Cours1BAC SVT1Pas encore d'évaluation
- COURS BC - PlasteDocument23 pagesCOURS BC - PlasteMohammed El MachmoulPas encore d'évaluation
- La Photosynthã Se Chez Les végétauxDocument39 pagesLa Photosynthã Se Chez Les végétauxBouchra OrPas encore d'évaluation
- Complexes Photosynthétiques 2Document2 pagesComplexes Photosynthétiques 2rizette10Pas encore d'évaluation
- Role Des Pigments Chlorophylliens Dans La Captation de Lenergie Lumineuse PDF 3Document19 pagesRole Des Pigments Chlorophylliens Dans La Captation de Lenergie Lumineuse PDF 3ekpaou eyoutchah mazamaPas encore d'évaluation
- CHAP1 - PV - Chap2 - Nutrition Carbonée Part1Document10 pagesCHAP1 - PV - Chap2 - Nutrition Carbonée Part1Sagacious Ivejuten100% (1)
- L'Absorption de La LumièreDocument8 pagesL'Absorption de La LumièreAyoub Abdessalem Djebbar100% (1)
- Métabolisme - 081427Document66 pagesMétabolisme - 081427Abel MalléPas encore d'évaluation
- Cours Chapitre 2Document13 pagesCours Chapitre 2carlos.h.farias00Pas encore d'évaluation
- Généralités Sur La PhotosynthèseDocument2 pagesGénéralités Sur La PhotosynthèseIlmi Guelleh0% (1)
- Production de La Matière Organique Et Flux D'énergieDocument22 pagesProduction de La Matière Organique Et Flux D'énergieViperDi AB100% (1)
- Chapitre 3 - Nutrition Carbonée - Prof Ayoub KASRATIDocument40 pagesChapitre 3 - Nutrition Carbonée - Prof Ayoub KASRATISomeone IdkPas encore d'évaluation
- Chapitre 2Document5 pagesChapitre 2Kendor94Pas encore d'évaluation
- Chap1 PhotosyntheseDocument2 pagesChap1 PhotosyntheseAbdoulaye Junior DialloPas encore d'évaluation
- Student-NUTRITION CARBONEE-2021-2022Document99 pagesStudent-NUTRITION CARBONEE-2021-2022IDTH EBM ApplicationPas encore d'évaluation
- Correction TP2Document3 pagesCorrection TP2Pilote Yamadjako100% (1)
- Chapitre 2Document25 pagesChapitre 2Mohamed OuanaimPas encore d'évaluation
- 1 10-Photosynthese PDFDocument66 pages1 10-Photosynthese PDFFitahianaPas encore d'évaluation
- Chap 4 Métabolisme FUE-SVT 2020-21.Ppt Partie 1Document56 pagesChap 4 Métabolisme FUE-SVT 2020-21.Ppt Partie 1Fatima zahrae AabidaPas encore d'évaluation
- Les Mécanismes Fondamentaux de La PhotosynthèseDocument24 pagesLes Mécanismes Fondamentaux de La PhotosynthèseJohns AtsacwounPas encore d'évaluation
- Ecosys MicrobDocument33 pagesEcosys MicrobRanya AchetPas encore d'évaluation
- Campbell, Réponses Cahier Coop ch4Document4 pagesCampbell, Réponses Cahier Coop ch4simmboisPas encore d'évaluation
- Chapitre 6, PhotosynthèseDocument22 pagesChapitre 6, PhotosynthèseAbdou BayePas encore d'évaluation
- Chapitre 10Document21 pagesChapitre 10agirmanmazlum3468Pas encore d'évaluation
- Nutrition Carbonée Et Approvisionnement Energétique - CompletDocument31 pagesNutrition Carbonée Et Approvisionnement Energétique - CompletYassmine AbramPas encore d'évaluation
- JL Ne1 Mod2 Exercice - 12 CORR A15Document2 pagesJL Ne1 Mod2 Exercice - 12 CORR A15Dyhia HmdPas encore d'évaluation
- Respiration Cellulaire PR Rajaonera TovoDocument46 pagesRespiration Cellulaire PR Rajaonera TovoFahendrenaPas encore d'évaluation
- PhotosynthèseDocument6 pagesPhotosynthèsemarceau100% (1)
- ChloroplasteDocument4 pagesChloroplasteHugo PerazziPas encore d'évaluation
- Biologie Végétale L1 SVT 2024Document79 pagesBiologie Végétale L1 SVT 2024Ay ManPas encore d'évaluation
- Comment Être Plus Éloquent - 15 Étapes (Avec Images)Document5 pagesComment Être Plus Éloquent - 15 Étapes (Avec Images)Ay Man100% (1)
- Bank DR Ongali 2021Document29 pagesBank DR Ongali 2021Ay ManPas encore d'évaluation
- 2-Fécondation Contraception AMPDocument49 pages2-Fécondation Contraception AMPAy ManPas encore d'évaluation
- 8 MalformationsDocument65 pages8 MalformationsAy ManPas encore d'évaluation
- 4-Le Tissu ConjonctifDocument60 pages4-Le Tissu ConjonctifAy ManPas encore d'évaluation
- 4-Période EmbryonnaireDocument125 pages4-Période EmbryonnaireAy ManPas encore d'évaluation
- Sujets D'examens D'anatomie Corrigés 2Document71 pagesSujets D'examens D'anatomie Corrigés 2Ay ManPas encore d'évaluation
- NEO ANATOMIE DE SYNTHESE TOME 6 VERSION L.M.D - CopieDocument38 pagesNEO ANATOMIE DE SYNTHESE TOME 6 VERSION L.M.D - CopieAy ManPas encore d'évaluation
- 6-Les Populations Cellulaires LibresDocument50 pages6-Les Populations Cellulaires LibresAy ManPas encore d'évaluation
- NEO PHYSIOLGIE TOM 5 RespiratoireDocument29 pagesNEO PHYSIOLGIE TOM 5 RespiratoireAy ManPas encore d'évaluation
- 2022 anatomie.L1.BM Système - Squelettique 12.01Document35 pages2022 anatomie.L1.BM Système - Squelettique 12.01Ay ManPas encore d'évaluation
- Appareil de GolgiDocument5 pagesAppareil de GolgiAy ManPas encore d'évaluation
- Cyto1an Mitochondrie2022aouatiDocument12 pagesCyto1an Mitochondrie2022aouatiAy ManPas encore d'évaluation
- Activité 1 Unicellulaires TPDocument2 pagesActivité 1 Unicellulaires TPefnt2023Pas encore d'évaluation
- Operation Dark WinterDocument1 pageOperation Dark WinterAnonymous 3X5KbLLiDjPas encore d'évaluation
- 1964 Vivre Avec Le Lama (Rampa)Document225 pages1964 Vivre Avec Le Lama (Rampa)alonso2016Pas encore d'évaluation
- GFA Section Et Groupes-3-4Document2 pagesGFA Section Et Groupes-3-4Dexter DexterPas encore d'évaluation
- 086 RouichiDocument12 pages086 RouichiHanane AzurePas encore d'évaluation
- Les ReinsDocument27 pagesLes ReinsDidi Hmitoche100% (2)
- Rapport Can 2016 PaysagisteDocument2 pagesRapport Can 2016 PaysagisteNZOMOPas encore d'évaluation
- تمارين مع الحل للمراجعة في اللغة الفرنسية للسنة الرابعة و الخامسة ابتدائي - الجيل الثانيDocument31 pagesتمارين مع الحل للمراجعة في اللغة الفرنسية للسنة الرابعة و الخامسة ابتدائي - الجيل الثانيرحيق الزهورPas encore d'évaluation
- TFC MakayaDocument57 pagesTFC MakayaBenito Benito100% (1)
- Sdic PL0005Document44 pagesSdic PL0005Ghizlane GhazalPas encore d'évaluation
- 4.ETUDE TECHNIK TomateDocument20 pages4.ETUDE TECHNIK TomateIsmail AlaouiPas encore d'évaluation
- Compte-Rendus Physiologie Animale PDFDocument48 pagesCompte-Rendus Physiologie Animale PDFHanane Jahid60% (5)
- Physio Cours SynthèseDocument127 pagesPhysio Cours SynthèsesamoyedePas encore d'évaluation
- Daniel Barthélémy Cecilia Brion Javier PuntieriDocument241 pagesDaniel Barthélémy Cecilia Brion Javier PuntieriCécile BerlanPas encore d'évaluation
- Thérapie CellulaireDocument13 pagesThérapie CellulaireAhmedPas encore d'évaluation
- Genetique FormellDocument4 pagesGenetique FormellChafi Driss100% (1)
- La Cellule CoursDocument11 pagesLa Cellule Courschaimaa chamouniPas encore d'évaluation
- Memoire LAVENANT AstridDocument67 pagesMemoire LAVENANT Astridastrid.lavenant.mkdePas encore d'évaluation
- La Découverte de L'homme de Florès: 2ème PARTIE - Exercice 2 - Génétique Et Évolution (5 Points)Document2 pagesLa Découverte de L'homme de Florès: 2ème PARTIE - Exercice 2 - Génétique Et Évolution (5 Points)Soumana SogobaPas encore d'évaluation
- Revision Reproduction Humaine CorrigeDocument29 pagesRevision Reproduction Humaine CorrigeDavid KingPas encore d'évaluation
- SALAMEH 2021 ArchivageDocument145 pagesSALAMEH 2021 ArchivageImene ImenePas encore d'évaluation
- Le Compostage PDFDocument8 pagesLe Compostage PDFTanguy Zgeg ArrigoniPas encore d'évaluation
- Item 18: Grossesse Extra-Utérine (GEU) (Évaluations)Document5 pagesItem 18: Grossesse Extra-Utérine (GEU) (Évaluations)outsiderPas encore d'évaluation
- Arnoldo Palacios La Foret Et La PluieDocument7 pagesArnoldo Palacios La Foret Et La PluieMarioPas encore d'évaluation
- Etude Sur Le Miel de NiaouliDocument103 pagesEtude Sur Le Miel de NiaouliMidouri DjafferPas encore d'évaluation
- François Denis Bactériologie Médicale PDFDocument615 pagesFrançois Denis Bactériologie Médicale PDFabdellah abdelkhalekPas encore d'évaluation