Vous aimerez peut-être aussi
- Muscles: Les Grands Articles d'UniversalisD'EverandMuscles: Les Grands Articles d'UniversalisPas encore d'évaluation
- Neurobiologie: Les Grands Articles d'UniversalisD'EverandNeurobiologie: Les Grands Articles d'UniversalisPas encore d'évaluation
- B-Physiologie de La Motricité SomatiqueDocument22 pagesB-Physiologie de La Motricité SomatiqueNoura AzaouagPas encore d'évaluation
- Correction Td2: Le Circuit Neuronique Du Réflexe de Flexion Chez La GrenouilleDocument2 pagesCorrection Td2: Le Circuit Neuronique Du Réflexe de Flexion Chez La GrenouillemicproteoPas encore d'évaluation
- CoursreflexemyotatiqueDocument7 pagesCoursreflexemyotatiqueAmor FaouziPas encore d'évaluation
- Cours Neuro Du 19 Et 20 Mars 2020Document4 pagesCours Neuro Du 19 Et 20 Mars 2020Abounasre khadihaPas encore d'évaluation
- SSV5U25C Perception à Laction 2020 L3 Neurosciences 2 Partie 2 Réflexes Spinaux Et Posture - TrouslardDocument32 pagesSSV5U25C Perception à Laction 2020 L3 Neurosciences 2 Partie 2 Réflexes Spinaux Et Posture - TrouslardMaddy IlPas encore d'évaluation
- IV. Le Récepteur de GolgiDocument107 pagesIV. Le Récepteur de GolgiAminou MounirouPas encore d'évaluation
- II Les Réflexes ProprioceptifsDocument31 pagesII Les Réflexes ProprioceptifsHamza Abu Humaid100% (2)
- 2 - Physiologie de La MotricitéDocument115 pages2 - Physiologie de La MotricitéBouchafra AbderahimPas encore d'évaluation
- SVT - Fiche 14 - RéflexesDocument8 pagesSVT - Fiche 14 - RéflexesClemence BrasseletPas encore d'évaluation
- SEQ 4-COURS Redige-Comportement-ReflexeDocument6 pagesSEQ 4-COURS Redige-Comportement-ReflexeLadetPas encore d'évaluation
- FA1 Correction-2Document9 pagesFA1 Correction-2Emmanuel MaxPas encore d'évaluation
- Sy Nerveux PrimaireDocument15 pagesSy Nerveux PrimairePark SanaPas encore d'évaluation
- Le Système Nerveux KHDocument12 pagesLe Système Nerveux KHayaezzouhri312Pas encore d'évaluation
- Anatomie 08Document3 pagesAnatomie 08Hamza Adri100% (1)
- Les Reflexes Cours BPA PDFDocument14 pagesLes Reflexes Cours BPA PDFKomi HoussounoukpePas encore d'évaluation
- Le Fonctionnement Du Systeme NerveuxDocument11 pagesLe Fonctionnement Du Systeme NerveuxSadi KoukouPas encore d'évaluation
- Réflexe de Flexion4eme Maths Version 1Document4 pagesRéflexe de Flexion4eme Maths Version 1Aziz BenamorPas encore d'évaluation
- 1-Physiologie Des Récepteurs Sensoriels PDFDocument34 pages1-Physiologie Des Récepteurs Sensoriels PDFImad Imad ZaroutaPas encore d'évaluation
- I) Evaluation Des Resssources Partie A: Evaluation Des Savoirs Exercice 1: Questions À Choix Multiples (QCM)Document8 pagesI) Evaluation Des Resssources Partie A: Evaluation Des Savoirs Exercice 1: Questions À Choix Multiples (QCM)mahopfranck092Pas encore d'évaluation
- Physiologie Du Système NerveuxDocument24 pagesPhysiologie Du Système NerveuxNa Yomi100% (1)
- Diagnostic DifferentielDocument4 pagesDiagnostic Differentielal nitakPas encore d'évaluation
- Nerfs Craniens (Achraf Mesfioui) PDFDocument2 pagesNerfs Craniens (Achraf Mesfioui) PDFAbir Saalaoui100% (1)
- Neurophysio de La FCT ProprioceptiveDocument8 pagesNeurophysio de La FCT ProprioceptiveImane BenderazPas encore d'évaluation
- Role Du Systeme Nerveux Dans Le Comportement Moteur Dun Animal - ts2 SunudaaraDocument10 pagesRole Du Systeme Nerveux Dans Le Comportement Moteur Dun Animal - ts2 Sunudaaradjibril diattaPas encore d'évaluation
- RachisDocument39 pagesRachisJuliette THIBAUTPas encore d'évaluation
- Physiologie Neuro-MuscuclaireDocument53 pagesPhysiologie Neuro-MuscuclaireMampionona RakotonandrasanaPas encore d'évaluation
- La Fonction MotriceDocument11 pagesLa Fonction Motricemohamed1539Pas encore d'évaluation
- Chap4 PapierDocument24 pagesChap4 PapierChristian abahPas encore d'évaluation
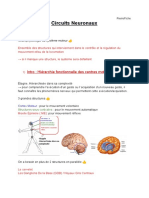
- Circuits NeuronauxDocument23 pagesCircuits NeuronauxPercheryPas encore d'évaluation
- UE 3 Reflexe MedullaireDocument6 pagesUE 3 Reflexe MedullaireCamille DruonPas encore d'évaluation
- La Neurophysiologie Cours 2013Document17 pagesLa Neurophysiologie Cours 2013friends diaryPas encore d'évaluation
- Les Activités Reflexes 2013Document6 pagesLes Activités Reflexes 2013Gnandi BilantéPas encore d'évaluation
- Techniques Ten Dine Uses Decordage Et VibrationsDocument7 pagesTechniques Ten Dine Uses Decordage Et VibrationsJomhod JomPas encore d'évaluation
- Boutonnat Jean p01 PDFDocument61 pagesBoutonnat Jean p01 PDFMoncef PechaPas encore d'évaluation
- Fiche Technique 7 Observation Du Nerf Dilacere Cellule NerveuseDocument2 pagesFiche Technique 7 Observation Du Nerf Dilacere Cellule NerveuseBenhsinat ChaimaaPas encore d'évaluation
- 2 Physiologie Reflexe Academic TeamDocument15 pages2 Physiologie Reflexe Academic TeamUubay 2003Pas encore d'évaluation
- Le Système Nerveu x22 22 Cours.Document8 pagesLe Système Nerveu x22 22 Cours.meryemPas encore d'évaluation
- Système Musculaire 2020Document22 pagesSystème Musculaire 2020Sarah OuajnatiPas encore d'évaluation
- Tp13 Circuit Neuronique Du Reflexe Achilleen Tres ImportantDocument3 pagesTp13 Circuit Neuronique Du Reflexe Achilleen Tres Importantibrahim abdillahiPas encore d'évaluation
- Devoir de Contrôle N°1 - SVT - Bac Mathématiques (2013-2014) MR TRIFI Mohamed TaharDocument3 pagesDevoir de Contrôle N°1 - SVT - Bac Mathématiques (2013-2014) MR TRIFI Mohamed TaharOussema BejiPas encore d'évaluation
- Anatomie FonctionnelleDocument44 pagesAnatomie FonctionnelleAbir SaalaouiPas encore d'évaluation
- SN Périphérique 1.0 2Document6 pagesSN Périphérique 1.0 2SayPas encore d'évaluation
- l3 Neurophysio Chap4Document16 pagesl3 Neurophysio Chap4nobodyPas encore d'évaluation
- Hornung - 6 Nerfs VaisseauDocument14 pagesHornung - 6 Nerfs Vaisseauyago.rodriguezcPas encore d'évaluation
- TP ReflexeDocument7 pagesTP ReflexebenjamingbodoPas encore d'évaluation
- Les Racines RachidiennesDocument10 pagesLes Racines RachidiennesMohamed Abdenour ChettouhPas encore d'évaluation
- NeuroDocument39 pagesNeuroClement / ZwiifyPas encore d'évaluation
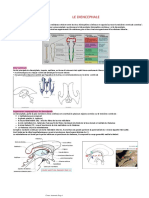
- SVT Le Reflexe Myotatique PDFDocument7 pagesSVT Le Reflexe Myotatique PDFAnonymous TrrvM8Pas encore d'évaluation
- Examen NeurologiqueDocument11 pagesExamen NeurologiqueHoussemPas encore d'évaluation
- 2023-3BB - Communication NerveuseDocument63 pages2023-3BB - Communication NerveuseIsabella AlmeidaPas encore d'évaluation
- Physiologie NerveuseDocument10 pagesPhysiologie NerveuseMouad HiliaPas encore d'évaluation
- Système Nerveux AutonomeDocument43 pagesSystème Nerveux AutonomeAbderrahmane SarPas encore d'évaluation
- Reflexe Myotatique PDFDocument13 pagesReflexe Myotatique PDFAnonymous iV3WIJnOxPas encore d'évaluation
- Nadia Cours 3AC 2éme Semestre CompletDocument53 pagesNadia Cours 3AC 2éme Semestre Completcrmefgroupesvt2Pas encore d'évaluation
- Nerf SciatiqueDocument5 pagesNerf SciatiqueYohann CdyPas encore d'évaluation
- 2-Résumé Lydia Tadjadit Part2 PDFDocument42 pages2-Résumé Lydia Tadjadit Part2 PDFSaibi100% (1)
- Membres: Les Grands Articles d'UniversalisD'EverandMembres: Les Grands Articles d'UniversalisPas encore d'évaluation
- Sommeil: Les Grands Articles d'UniversalisD'EverandSommeil: Les Grands Articles d'UniversalisPas encore d'évaluation
- Le Système NerveuxDocument8 pagesLe Système NerveuxazizPas encore d'évaluation
- SD CerebDocument22 pagesSD Cereb31243564Pas encore d'évaluation
- Correction Quiz 2Document9 pagesCorrection Quiz 2chamsedinerourPas encore d'évaluation
- Exercices D'harmonisation Pour Décoder Les Émotions, Pacifier Le Corps Et Prévenir Les Maladies PDFDocument184 pagesExercices D'harmonisation Pour Décoder Les Émotions, Pacifier Le Corps Et Prévenir Les Maladies PDFLaboratoire Saïdi100% (1)
- Generalite Sur La GymnastiqueDocument6 pagesGeneralite Sur La GymnastiqueRomulus FARARA100% (1)
- Néphrectomie Simple Et Néphrectomie Elargie Pour CancerDocument28 pagesNéphrectomie Simple Et Néphrectomie Elargie Pour Cancerdjasmi mohamedPas encore d'évaluation
- Manifestations Ophtalmiques Orbitopalpebrales de L'obstruction Nasale. Di R. Gola Et Al., 13 09 15Document14 pagesManifestations Ophtalmiques Orbitopalpebrales de L'obstruction Nasale. Di R. Gola Et Al., 13 09 15Anders CroccoPas encore d'évaluation
- Séquence 3 Tle DDocument8 pagesSéquence 3 Tle DtomeyumPas encore d'évaluation
- Luxations Du CoudeDocument3 pagesLuxations Du CoudeNouaïri Amine100% (1)
- Imagerie Anatomique Des Nerfs CraniensDocument12 pagesImagerie Anatomique Des Nerfs Craniensf.ramondencPas encore d'évaluation
- 13 Articulation Du CoudeDocument66 pages13 Articulation Du CoudeLamia LamiaPas encore d'évaluation
- PR Bouziane - Anatomie Chirurgicale de L'estomac - CM PDFDocument70 pagesPR Bouziane - Anatomie Chirurgicale de L'estomac - CM PDFmomoPas encore d'évaluation
- Les Os CoxauxDocument8 pagesLes Os CoxauxSylvain GravelaisPas encore d'évaluation
- Diencéphale by Imy BensDocument3 pagesDiencéphale by Imy BensIMENELEPas encore d'évaluation
- Systeme Nerveux AutonomeDocument35 pagesSysteme Nerveux AutonomeSadi Koukou100% (1)
- Atlas D'anatomie Des Membres - Nerfs Peripheriques by Philippe Rigoard PDFDocument340 pagesAtlas D'anatomie Des Membres - Nerfs Peripheriques by Philippe Rigoard PDFPedro Miiguel100% (1)
- Cours - Respiration Et Circulation SanguineDocument6 pagesCours - Respiration Et Circulation SanguineHicham Elamrani100% (1)
- Lithiase SalivaireDocument18 pagesLithiase SalivaireMustfa YaseenPas encore d'évaluation
- Le Système Musculosquelettique DébutDocument62 pagesLe Système Musculosquelettique DébutCheryl MacdonaldPas encore d'évaluation
- Anatomie Du LarynxDocument8 pagesAnatomie Du LarynxPonlue StephenPas encore d'évaluation
- 8 Les Articulateurs Er Arcs Faciaux 2022.Document7 pages8 Les Articulateurs Er Arcs Faciaux 2022.Christ stPas encore d'évaluation
- UntitledDocument201 pagesUntitledabPas encore d'évaluation
- Anatomie Chirurgicale Du PelvisDocument30 pagesAnatomie Chirurgicale Du Pelvisdjasmi mohamedPas encore d'évaluation
- 2009 Traitement Par Reconstruction Posturale Dune Gonarthrose Bilaterale Sur Genu Varum A Propos Dun Cas PDFDocument100 pages2009 Traitement Par Reconstruction Posturale Dune Gonarthrose Bilaterale Sur Genu Varum A Propos Dun Cas PDFMjidKarim0% (1)
- Fracture de La Palette Humérale de L - Enfant - CopieDocument4 pagesFracture de La Palette Humérale de L - Enfant - Copieikou ikramPas encore d'évaluation
- Peat Set 1 Examen Pratique SOULIGNÉDocument4 pagesPeat Set 1 Examen Pratique SOULIGNÉScribdTranslationsPas encore d'évaluation
- Memoir de Fin D'etudes - La Prise en Charge Et Appareillage D'un Ampute FemoralDocument48 pagesMemoir de Fin D'etudes - La Prise en Charge Et Appareillage D'un Ampute Femoralseddik chanaouiPas encore d'évaluation
- Programme Detente Sans Materiel-2Document53 pagesProgramme Detente Sans Materiel-2Spécialiste HardwarePas encore d'évaluation
- 1-Généralité Sur Le Corps HumainDocument6 pages1-Généralité Sur Le Corps HumaindjoualalaePas encore d'évaluation
- Voies D Abord de L Aorte AbdominaleDocument23 pagesVoies D Abord de L Aorte AbdominaleM. AdnenePas encore d'évaluation