Vous aimerez peut-être aussi
- Faculte de Medecine de Nouakchott: Immunologie FondamentaleDocument211 pagesFaculte de Medecine de Nouakchott: Immunologie FondamentaleSaadhah HahPas encore d'évaluation
- Le Système ImmunitaireDocument18 pagesLe Système ImmunitaireccPas encore d'évaluation
- Le Système ImmunitaireDocument9 pagesLe Système ImmunitairendiayePas encore d'évaluation
- Immunologie II (Suite 2)Document52 pagesImmunologie II (Suite 2)مصطفى الورديغيPas encore d'évaluation
- Cours ImmunologieDocument285 pagesCours ImmunologiedouinienPas encore d'évaluation
- Vue Générale Du Système ImmunitaireDocument20 pagesVue Générale Du Système ImmunitaireAbdelhamid ArouiPas encore d'évaluation
- ImmunologieDocument24 pagesImmunologieayoub berrePas encore d'évaluation
- Vue Générale Du Système ImmunitaireDocument18 pagesVue Générale Du Système ImmunitairedcdPas encore d'évaluation
- Immunologie IDocument107 pagesImmunologie Iمصطفى الورديغيPas encore d'évaluation
- Immunologie 1Document52 pagesImmunologie 1Mustafa KouzritePas encore d'évaluation
- L'Immunologie - SVT 1èrea Tome2Document7 pagesL'Immunologie - SVT 1èrea Tome2Guy TangaPas encore d'évaluation
- Le Systeme ImmunitaireDocument32 pagesLe Systeme ImmunitaireIlyes MraidiaPas encore d'évaluation
- Cours 1 Immunologie - Généralités-ConvertiDocument38 pagesCours 1 Immunologie - Généralités-Convertinenend33Pas encore d'évaluation
- L'Immunité SpécifiqueDocument12 pagesL'Immunité Spécifiqueraslenbouallegue12345Pas encore d'évaluation
- ImmunoDocument29 pagesImmunoIkram DahmaniPas encore d'évaluation
- Système immunitaire: Les Grands Articles d'UniversalisD'EverandSystème immunitaire: Les Grands Articles d'UniversalisPas encore d'évaluation
- S2 Immunologie Khither Et MadouiDocument59 pagesS2 Immunologie Khither Et MadouiAmina AitPas encore d'évaluation
- Cours Immunologie KHITHERDocument97 pagesCours Immunologie KHITHEROumia Harbit100% (2)
- Immunologie ExposeDocument34 pagesImmunologie ExposeSafia Amahmid100% (11)
- Immuno 1Document13 pagesImmuno 1mouadennaji519Pas encore d'évaluation
- Les Réponses Immunitaires DR MILOUDIDocument13 pagesLes Réponses Immunitaires DR MILOUDIFarah B. BtoushPas encore d'évaluation
- 97 1bDocument8 pages97 1bhizballah27Pas encore d'évaluation
- FR - Semaine 1 - FinalDocument43 pagesFR - Semaine 1 - FinalRima OswaldPas encore d'évaluation
- Cours Immunologie S5Document113 pagesCours Immunologie S5hamzaabzou154Pas encore d'évaluation
- Bios 308 Immunologie Generale Et FonctionelleDocument159 pagesBios 308 Immunologie Generale Et FonctionelleMahamat Mahamat AboubakarPas encore d'évaluation
- ImmunologieDocument28 pagesImmunologieImenePas encore d'évaluation
- Anapathbest - Nouvele VersionDocument39 pagesAnapathbest - Nouvele VersionnathankambPas encore d'évaluation
- Resumé Du Cours de L'immunitéDocument3 pagesResumé Du Cours de L'immunitéMomme Ould Deyahi Achrif100% (7)
- TD ImmDocument35 pagesTD ImmWiem BenchahbounPas encore d'évaluation
- Cours ImmunoDocument29 pagesCours ImmunoChristian NgalaPas encore d'évaluation
- Cours IV Tolérance Immunitaire Et Auto ImmunitéDocument13 pagesCours IV Tolérance Immunitaire Et Auto ImmunitétinaPas encore d'évaluation
- Chapitre 1. Introduction Au Système ImmunitaireDocument69 pagesChapitre 1. Introduction Au Système Immunitairefallone zannouPas encore d'évaluation
- Introduction 2021-2022 PDFDocument75 pagesIntroduction 2021-2022 PDFStephanePas encore d'évaluation
- Fiche IntroDocument3 pagesFiche IntrorequiemforathrowawayPas encore d'évaluation
- RappelDocument11 pagesRappelNadjib A. NedjarPas encore d'évaluation
- Chapitre 1 Introduction À L'immunologieDocument83 pagesChapitre 1 Introduction À L'immunologieHayat IssaouiPas encore d'évaluation
- Radio-Immuno Assays (Ria) o l2Document45 pagesRadio-Immuno Assays (Ria) o l2EngwataPas encore d'évaluation
- 2eme Biotech 19-20 Immunologie Générale AllouniDocument67 pages2eme Biotech 19-20 Immunologie Générale AllouniMaï Ssa100% (1)
- CHAPITRE 4 Immunité InnéeDocument13 pagesCHAPITRE 4 Immunité InnéeJuan Sebastian MoraPas encore d'évaluation
- Cours 1 d'Immuno2LSP23Document27 pagesCours 1 d'Immuno2LSP23abdou2004trPas encore d'évaluation
- FR - Semaine 1 - Final PDFDocument43 pagesFR - Semaine 1 - Final PDFChristian GaudinPas encore d'évaluation
- M1 Im 19-20 Sobhi W M1 Imm Cours de Bet DDocument47 pagesM1 Im 19-20 Sobhi W M1 Imm Cours de Bet DBio Bioch100% (1)
- 32 Chap 26Document13 pages32 Chap 26Fadila MararPas encore d'évaluation
- 1 Er Cours Immuno Appl BS 202 PDFDocument10 pages1 Er Cours Immuno Appl BS 202 PDFHasna BentPas encore d'évaluation
- Cours Immunologie - 2ème Sciences Biologiques Section 01 - OUABEDDocument19 pagesCours Immunologie - 2ème Sciences Biologiques Section 01 - OUABEDDjihadnoorPas encore d'évaluation
- Cours Immunologie - 2ème Sciences Biologiques Section 01 - OUABEDDocument22 pagesCours Immunologie - 2ème Sciences Biologiques Section 01 - OUABEDKakashi HatakéPas encore d'évaluation
- Suite Cours D - Immunologie PDFDocument61 pagesSuite Cours D - Immunologie PDFfafi Ali100% (1)
- Introduction ImmunologieDocument29 pagesIntroduction Immunologieagag.salahPas encore d'évaluation
- Myasthénie Auto-Immune: Dr. L.Larbi Cherif Hmusora/1°RmDocument92 pagesMyasthénie Auto-Immune: Dr. L.Larbi Cherif Hmusora/1°RmHakimoPas encore d'évaluation
- INTRODUCTION A L'immunologieDocument21 pagesINTRODUCTION A L'immunologieRodrigue AssohPas encore d'évaluation
- La Réponse Immunitaire Innée Et Adaptative ModifDocument8 pagesLa Réponse Immunitaire Innée Et Adaptative ModifAni TrghwiPas encore d'évaluation
- Introduction Au Système ImmunitaireDocument47 pagesIntroduction Au Système ImmunitaireEmmanuel wendkuni BagrePas encore d'évaluation
- ImmunoDocument5 pagesImmunoAmadou CISSEPas encore d'évaluation
- Cours 1Document71 pagesCours 1midoan100% (1)
- Physiopathologie Mai Final 2Document5 pagesPhysiopathologie Mai Final 2EbePas encore d'évaluation
- La Reconnaissance Du Soi Et Du Non SoiDocument6 pagesLa Reconnaissance Du Soi Et Du Non SoiRonan PerennesPas encore d'évaluation
- Cours Apc TC Defense de Organisme Fiche ÉlèveDocument23 pagesCours Apc TC Defense de Organisme Fiche ÉlèveDAOUDA Nable CoulibalyPas encore d'évaluation
- Biologie 2Document57 pagesBiologie 2karen StéphaniePas encore d'évaluation
- Introduction A L'immunologieDocument28 pagesIntroduction A L'immunologiemelchiPas encore d'évaluation
- Cahier 49 - Viellissement Hormonal-Tome 2Document94 pagesCahier 49 - Viellissement Hormonal-Tome 2Adi MaghiarPas encore d'évaluation
- Cahier 16 - Immuno-Allergie (Tome 2)Document73 pagesCahier 16 - Immuno-Allergie (Tome 2)Adi MaghiarPas encore d'évaluation
- Dematteis Maurice P01Document14 pagesDematteis Maurice P01Adi MaghiarPas encore d'évaluation
- Complexul HLADocument75 pagesComplexul HLAAdi MaghiarPas encore d'évaluation
- Les Bases Théoriques Des Groupes Sanguins ABO, RH, KellDocument20 pagesLes Bases Théoriques Des Groupes Sanguins ABO, RH, KellAdi MaghiarPas encore d'évaluation
- Les Bases Théoriques Des Groupes Sanguins ABO, RH, KellDocument20 pagesLes Bases Théoriques Des Groupes Sanguins ABO, RH, KellAdi MaghiarPas encore d'évaluation
- S SVT Obligatoire 2017 Centres Etrangers CorrigeDocument9 pagesS SVT Obligatoire 2017 Centres Etrangers CorrigeWilfriedPas encore d'évaluation
- Bac Blanc 2017 FinaleDocument4 pagesBac Blanc 2017 FinaleAmor FaouziPas encore d'évaluation
- Pakao SVT TleDocument220 pagesPakao SVT TleLĘFÂPas encore d'évaluation
- 1notions Du Soi Et Du Non Soi CoursDocument6 pages1notions Du Soi Et Du Non Soi CoursAnouar NACERIPas encore d'évaluation
- Ganglion LymphatiqueDocument6 pagesGanglion LymphatiqueZeroualPas encore d'évaluation
- Brevet 1Document2 pagesBrevet 1bledburstPas encore d'évaluation
- FrenchJournal 26 ViralDocument24 pagesFrenchJournal 26 ViralprincessePas encore d'évaluation
- RMS 579 1797Document6 pagesRMS 579 1797Sarab AlwardPas encore d'évaluation
- Immunologie Exercices Corriges 1Document3 pagesImmunologie Exercices Corriges 1sonilola80% (5)
- Fiche de TD Sur La Reponse ImmunitaireDocument5 pagesFiche de TD Sur La Reponse ImmunitaireLONGTOUO JaurèsPas encore d'évaluation
- Immuno QCMDocument2 pagesImmuno QCMDoc Zak91% (11)
- Infections Neonatales em - 2Document17 pagesInfections Neonatales em - 2IdiAmadouPas encore d'évaluation
- Simulation QCM Hé Matologie Biologique. 2AP. FMPR. PR Masrar 2022-2023 Version Corrigã© eDocument23 pagesSimulation QCM Hé Matologie Biologique. 2AP. FMPR. PR Masrar 2022-2023 Version Corrigã© eSami BouzbibaPas encore d'évaluation
- Immuno Td02Document2 pagesImmuno Td02Fatïma ZohraPas encore d'évaluation
- NIGELLEDocument7 pagesNIGELLEibrahimPas encore d'évaluation
- Memoire DjouadDocument123 pagesMemoire DjouadKouinine MinifootPas encore d'évaluation
- Immuno27 Hypersensibilite2Document28 pagesImmuno27 Hypersensibilite2Kanneaufii KaramelPas encore d'évaluation
- Cours de SVT PS Chapitre 19-Se Soigner Grâce Au Système ImmunitaireDocument4 pagesCours de SVT PS Chapitre 19-Se Soigner Grâce Au Système ImmunitaireOndzePas encore d'évaluation
- Ensp/Dz 2021-2022Document5 pagesEnsp/Dz 2021-2022AbdourahamanePas encore d'évaluation
- QCM Hemato CellulaireDocument4 pagesQCM Hemato CellulaireSouhayl Chouaaib100% (1)
- Livre ImmunologieDocument907 pagesLivre ImmunologieColin ManonPas encore d'évaluation
- Les Anticorps Sont Des Molécules Majeures de Notre OrganismeDocument2 pagesLes Anticorps Sont Des Molécules Majeures de Notre OrganismelilyaPas encore d'évaluation
- Diabete de Type 1 Une Maladie Auto Immune, Vraiment 2019Document8 pagesDiabete de Type 1 Une Maladie Auto Immune, Vraiment 2019FérielPas encore d'évaluation
- TD Immunologie Samari HDocument10 pagesTD Immunologie Samari Hمهدي دوشة محمدPas encore d'évaluation
- Immunotech ch1Document6 pagesImmunotech ch1atyoffice.dzPas encore d'évaluation
- Bio JuilletDocument124 pagesBio JuilletmariaPas encore d'évaluation
- SVT - Livret-corriges-Partie-02 PDFDocument19 pagesSVT - Livret-corriges-Partie-02 PDFinterPas encore d'évaluation
- Micro Immuno Therapie LucBodinDocument4 pagesMicro Immuno Therapie LucBodinramaflorePas encore d'évaluation
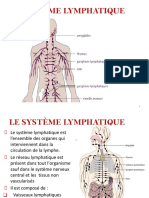
- 8-Le Système LymphatiqueDocument12 pages8-Le Système LymphatiqueImene ImenePas encore d'évaluation
- Réalisé Par: Supervisé ParDocument8 pagesRéalisé Par: Supervisé ParRomaissa KahlouchPas encore d'évaluation