Vous aimerez peut-être aussi
- La Dengue: Symptômes, traitement, prévention, où en est-on ?D'EverandLa Dengue: Symptômes, traitement, prévention, où en est-on ?Pas encore d'évaluation
- En 64c773234ebd6 Vol 50 No 28-English 1Document4 pagesEn 64c773234ebd6 Vol 50 No 28-English 1Lagath WanigabaduPas encore d'évaluation
- Les Grandes EndémiesDocument6 pagesLes Grandes EndémieskgtdPas encore d'évaluation
- Dengue FRDocument8 pagesDengue FRReinaldo AbentePas encore d'évaluation
- La DengueDocument3 pagesLa DengueDominique Da SilveiraPas encore d'évaluation
- Faire Face À L'apparition de Maladies Virales Infectieuses, Un Défi ContemporainDocument5 pagesFaire Face À L'apparition de Maladies Virales Infectieuses, Un Défi ContemporainBakolonPas encore d'évaluation
- Prevalence Et Evolution de La DengueDocument9 pagesPrevalence Et Evolution de La Denguehossein kouahiPas encore d'évaluation
- Makalah AntropologiDocument20 pagesMakalah AntropologiNYONGKERPas encore d'évaluation
- Entomologie Médicale Et Vétérinaire: Ouvrages / Book ReviewDocument2 pagesEntomologie Médicale Et Vétérinaire: Ouvrages / Book Reviewworld study portalsPas encore d'évaluation
- Ad 981218Document7 pagesAd 981218Dr Souleymane SackoPas encore d'évaluation
- Delatte 2008Document11 pagesDelatte 2008RaffaharianggaraPas encore d'évaluation
- 797-Article Text-1340-1-10-20200505 - FRDocument8 pages797-Article Text-1340-1-10-20200505 - FRBakolonPas encore d'évaluation
- 12-Dengue - Aspects Cliniques (Non Vu)Document5 pages12-Dengue - Aspects Cliniques (Non Vu)MONTOKI NINAPas encore d'évaluation
- Présentation CH 4Document22 pagesPrésentation CH 4hidayatab617Pas encore d'évaluation
- Fièvre Jaune, Épidémiologie Et Prévention Vaccinale Du VoyageurDocument3 pagesFièvre Jaune, Épidémiologie Et Prévention Vaccinale Du VoyageurAYOUB MOUHADDABPas encore d'évaluation
- Debarre-Gandon 2008Document28 pagesDebarre-Gandon 2008BourbakiPas encore d'évaluation
- Pathologie ÉmergenteDocument6 pagesPathologie ÉmergentekgtdPas encore d'évaluation
- Makalah Asuhan Keperawatan Pada Pasien Dengan Gangguan Sistem Imun Dan Hematologi: DHFDocument24 pagesMakalah Asuhan Keperawatan Pada Pasien Dengan Gangguan Sistem Imun Dan Hematologi: DHFMaria taebenuPas encore d'évaluation
- FRM DP Maladies InfectieusesDocument17 pagesFRM DP Maladies InfectieusesPhy RdaousPas encore d'évaluation
- Shigella: Un Agent Pathogène Hautement Virulent Et InsaisissableDocument7 pagesShigella: Un Agent Pathogène Hautement Virulent Et InsaisissableGuillame SawadogoPas encore d'évaluation
- Les Plus Grandes Épidémies Et Pandémies de L'histoire Cas de L'Algérie Entre L'époque Ottomane Et ColonialeDocument12 pagesLes Plus Grandes Épidémies Et Pandémies de L'histoire Cas de L'Algérie Entre L'époque Ottomane Et Colonialeka tiaPas encore d'évaluation
- Infections Fongiques en RéanimationDocument11 pagesInfections Fongiques en RéanimationSoumeya BelhimerPas encore d'évaluation
- 057-061 Variabilité Des Maladies Diarrhéiques Chez Les Enfants de Moins de 5 Ans en Milieu Urbain L'exemple de RufisquDocument5 pages057-061 Variabilité Des Maladies Diarrhéiques Chez Les Enfants de Moins de 5 Ans en Milieu Urbain L'exemple de RufisquChristelle NyadjouPas encore d'évaluation
- Chalghaf Bilel Doctel 2017 PDFADocument105 pagesChalghaf Bilel Doctel 2017 PDFAWilly ValePas encore d'évaluation
- Géographie Médicale Et Histoire Des CivilisationsDocument28 pagesGéographie Médicale Et Histoire Des CivilisationsMarcia SiqueiraPas encore d'évaluation
- En Afrique de L'ouest Et Du Centre: EpidémiesDocument28 pagesEn Afrique de L'ouest Et Du Centre: EpidémiesOjong YvesPas encore d'évaluation
- Article 3 Mevaa Dominique Vol 3 2 20161Document17 pagesArticle 3 Mevaa Dominique Vol 3 2 20161ZOUGA KOUNA Alain FlorentinPas encore d'évaluation
- TIC Cours Sept 2021Document59 pagesTIC Cours Sept 2021Zbreh SkeeperPas encore d'évaluation
- 22 2018 RisaDocument6 pages22 2018 RisaAmétépé HOUNMAVOPas encore d'évaluation
- ArbovirosesDocument14 pagesArbovirosesPellePas encore d'évaluation
- Fgioytr - CopieDocument462 pagesFgioytr - CopieOusmane DiakitéPas encore d'évaluation
- Virus de La DengueDocument46 pagesVirus de La DengueOuattara BamoryPas encore d'évaluation
- Analyse Politique COVİD-19Document7 pagesAnalyse Politique COVİD-19CGPas encore d'évaluation
- Trape 2011Document8 pagesTrape 2011diaibraPas encore d'évaluation
- Activité 3Document4 pagesActivité 3Kadiatou BarryPas encore d'évaluation
- 28 n0v 2021 Saadi Epidemiologie Des MTDocument31 pages28 n0v 2021 Saadi Epidemiologie Des MTsanaa.gamraniPas encore d'évaluation
- ÉpidémiologiesDocument85 pagesÉpidémiologiesOrianne AmoussouPas encore d'évaluation
- CoronaviruspourlesnulsDocument8 pagesCoronaviruspourlesnulsZahira SafouanPas encore d'évaluation
- Covid 19 Anglais NewDocument10 pagesCovid 19 Anglais NewLaurent Stefan SomePas encore d'évaluation
- Quae 36190Document27 pagesQuae 36190rahmahzemi58Pas encore d'évaluation
- Orora Et AlDocument9 pagesOrora Et Alororac_nyamoriPas encore d'évaluation
- Le Reseau de Distribution Deau Potable Un EcosystDocument38 pagesLe Reseau de Distribution Deau Potable Un EcosystIkram SebbaniPas encore d'évaluation
- 3 - Hygiene Hospitalière Et Infection NosocomialeDocument15 pages3 - Hygiene Hospitalière Et Infection NosocomialeomerPas encore d'évaluation
- 1les ZoonosesDocument8 pages1les ZoonosesEl-nabaoui ChaimaaPas encore d'évaluation
- Potential Health Risk Associated With Exposure To Microorganism in Urban FloodDocument4 pagesPotential Health Risk Associated With Exposure To Microorganism in Urban FloodRamesh SoniPas encore d'évaluation
- Maladie À Déclaration Obligatoire La Tuberculose en PTTDocument56 pagesMaladie À Déclaration Obligatoire La Tuberculose en PTTimene hammadiPas encore d'évaluation
- Maladies ÉmergentesDocument8 pagesMaladies ÉmergentesHenada Rania Lina IkramPas encore d'évaluation
- RMB32N3 DrepanocytoseDocument7 pagesRMB32N3 Drepanocytosesarahloba100Pas encore d'évaluation
- Épidémiologie Et Maladies Transmissibles 1Document43 pagesÉpidémiologie Et Maladies Transmissibles 1meryem erPas encore d'évaluation
- 002+Ao+Kone+Morbi Mortalite+Malinf+BouakeDocument7 pages002+Ao+Kone+Morbi Mortalite+Malinf+BouakeJpr AdjamePas encore d'évaluation
- Extrait Atlas ImmunologieDocument17 pagesExtrait Atlas ImmunologieDaniel NdPas encore d'évaluation
- Diakitel2syllab VihsidaDocument8 pagesDiakitel2syllab Vihsidaeric ouattaraPas encore d'évaluation
- 3-Infections Nosocomiales À Pseudomonas Aeruginosa Article 444444444444 PDFDocument4 pages3-Infections Nosocomiales À Pseudomonas Aeruginosa Article 444444444444 PDFRiadh HamdiPas encore d'évaluation
- Mazza Stalder2012Document13 pagesMazza Stalder2012NGATSONGO DIRAT Unis HaroldPas encore d'évaluation
- BrucelloseDocument10 pagesBrucelloseNoureddine SoltaniPas encore d'évaluation
- 2Document19 pages2Fatma HfdPas encore d'évaluation
- Les Problèmes de La SantéDocument3 pagesLes Problèmes de La SantéАлександра ОднолетковаPas encore d'évaluation
- Généralités Sur Les MDocument22 pagesGénéralités Sur Les MPabloPas encore d'évaluation
- Ankylostomes Et Ankylostomiase HumaineDocument5 pagesAnkylostomes Et Ankylostomiase HumaineentredosmundosPas encore d'évaluation
- Epidemiologie6an-12epidemiologie Maladies TransmissiblesDocument10 pagesEpidemiologie6an-12epidemiologie Maladies TransmissiblesTak YazPas encore d'évaluation
- SPHENOÏDEDocument24 pagesSPHENOÏDEGodwin BonkoungouPas encore d'évaluation
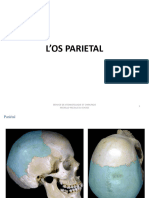
- Os ParietalDocument13 pagesOs ParietalGodwin BonkoungouPas encore d'évaluation
- 1 Muscles de L'epaule 2019Document45 pages1 Muscles de L'epaule 2019Godwin BonkoungouPas encore d'évaluation
- ExosDePhysInssa 2023Document8 pagesExosDePhysInssa 2023Godwin BonkoungouPas encore d'évaluation
- Exercice de Biochimie 2009-2010Document14 pagesExercice de Biochimie 2009-2010Godwin BonkoungouPas encore d'évaluation
- 3 Partie Caractéristiques Des LubrifiantDocument32 pages3 Partie Caractéristiques Des Lubrifiantsamir belamriPas encore d'évaluation
- Racines Carrees BaseDocument8 pagesRacines Carrees Basejulien9562Pas encore d'évaluation
- Mesure de Compression Moteur Vers FinaleDocument46 pagesMesure de Compression Moteur Vers FinaleRaouf HarzallahPas encore d'évaluation
- Jadwal Genap 2223-2Document2 pagesJadwal Genap 2223-2nowo benyPas encore d'évaluation
- Thèse Data IntegrityDocument83 pagesThèse Data IntegrityBasma YagoubiPas encore d'évaluation
- RSE & EthiqueDocument6 pagesRSE & Ethiquealemor2369Pas encore d'évaluation
- The Cuban Missile CrisisDocument8 pagesThe Cuban Missile Crisismilan.bodis523Pas encore d'évaluation
- Cahier D Exercices Ile Aux Mots 8hDocument88 pagesCahier D Exercices Ile Aux Mots 8hCizPas encore d'évaluation
- Exposé MDE Et Énergie RenouvelablesDocument12 pagesExposé MDE Et Énergie Renouvelablesromain fokamPas encore d'évaluation
- Les Étapes de Formation Des Roches SédimentaireDocument2 pagesLes Étapes de Formation Des Roches Sédimentairehamada2002Pas encore d'évaluation
- Caplp Externe Genie Electrique Electrotechnique Et Energie Epreuve 1 Doc RessourcesDocument28 pagesCaplp Externe Genie Electrique Electrotechnique Et Energie Epreuve 1 Doc RessourcesOus SàmàPas encore d'évaluation
- PV Liste-De-Prix Onduleurs HUAWEI 11-2023 FRDocument2 pagesPV Liste-De-Prix Onduleurs HUAWEI 11-2023 FRkoumbounisdimPas encore d'évaluation
- ExcisionDocument54 pagesExcisionAbdou Razak OuédraogoPas encore d'évaluation
- Canalisations de Gaz NaturelDocument120 pagesCanalisations de Gaz NaturelJean-David DelordPas encore d'évaluation
- Calendrier Des Examens Semestre Impair Janvier 2022 AlphaDocument28 pagesCalendrier Des Examens Semestre Impair Janvier 2022 AlphaMeg JustMegPas encore d'évaluation
- 3 Branches Triphasées en Injection Directe M215 Ou M250Document1 page3 Branches Triphasées en Injection Directe M215 Ou M250MbgardPas encore d'évaluation
- Observatoire National de La Filiere Riz Du Burkina Faso (Onriz)Document6 pagesObservatoire National de La Filiere Riz Du Burkina Faso (Onriz)toni_yousf2418Pas encore d'évaluation
- Introduction À La RobotiqueDocument19 pagesIntroduction À La RobotiqueRazzougui SarahPas encore d'évaluation
- Exercice D'application Optique VDocument1 pageExercice D'application Optique VARDALAn MohamedPas encore d'évaluation
- Controle Et Suivi Chantier RoutierhjhDocument14 pagesControle Et Suivi Chantier Routierhjhعثمان البريشيPas encore d'évaluation
- Exam. F.CDocument2 pagesExam. F.CmidsmasherPas encore d'évaluation
- 2nd - Exercices Corrigés - Variations D'une FonctDocument1 page2nd - Exercices Corrigés - Variations D'une Fonctalyahmed610Pas encore d'évaluation
- Analyse D'une Situation de Communication en TaDocument2 pagesAnalyse D'une Situation de Communication en Taroger martin bassong batiigPas encore d'évaluation
- Chapitre 1 LES OUTILS MATHEMATIQUESDocument9 pagesChapitre 1 LES OUTILS MATHEMATIQUESa.ddPas encore d'évaluation
- Process AciérieDocument6 pagesProcess Aciériesanae jaouiPas encore d'évaluation
- TFE Gustave KISHATU MWAMBA Version Finale-1Document112 pagesTFE Gustave KISHATU MWAMBA Version Finale-1gustave kishatu100% (2)
- Correction DevoirDeSyntheseN3 1AS 2009 FinDocument8 pagesCorrection DevoirDeSyntheseN3 1AS 2009 FinMehdi Ben Arif67% (3)
- AnnexeDocument168 pagesAnnexeMoez AliPas encore d'évaluation
- Memoire Inj Messaoud BENZOUAIDocument168 pagesMemoire Inj Messaoud BENZOUAIManong ShegueyPas encore d'évaluation
- Format Eur FrancaiseDocument1 pageFormat Eur FrancaiseAdnan NandaPas encore d'évaluation
- 20 Véritables remèdes de nos grands-mères pour maigrir vite et enfin perdre du poidsD'Everand20 Véritables remèdes de nos grands-mères pour maigrir vite et enfin perdre du poidsÉvaluation : 5 sur 5 étoiles5/5 (1)
- Améliorer votre mémoire: Un Guide pour l'augmentation de la puissance du cerveau, utilisant des techniques et méthodesD'EverandAméliorer votre mémoire: Un Guide pour l'augmentation de la puissance du cerveau, utilisant des techniques et méthodesÉvaluation : 5 sur 5 étoiles5/5 (2)
- La vie des abeilles: Prix Nobel de littératureD'EverandLa vie des abeilles: Prix Nobel de littératureÉvaluation : 4 sur 5 étoiles4/5 (41)
- Manuel pour les débutants Fabriquez des savons naturelsD'EverandManuel pour les débutants Fabriquez des savons naturelsÉvaluation : 3 sur 5 étoiles3/5 (2)
- LE PLACEBO, C'EST VOUS !: Comment donner le pouvoir à votre espritD'EverandLE PLACEBO, C'EST VOUS !: Comment donner le pouvoir à votre espritPas encore d'évaluation
- Régime Charge Glycémique, Régime Express IG: Perdre 10 kg et vivre 10 ans de plus ! + 39 Recettes IG basD'EverandRégime Charge Glycémique, Régime Express IG: Perdre 10 kg et vivre 10 ans de plus ! + 39 Recettes IG basÉvaluation : 5 sur 5 étoiles5/5 (3)
- Géologie de l'Amérique: Les Grands Articles d'UniversalisD'EverandGéologie de l'Amérique: Les Grands Articles d'UniversalisPas encore d'évaluation
- Beauté Santé: 107 Recettes faciles de produits cosmétiques bio à faire vous-mêmes avec les ingrédients de votre cuisine !D'EverandBeauté Santé: 107 Recettes faciles de produits cosmétiques bio à faire vous-mêmes avec les ingrédients de votre cuisine !Évaluation : 5 sur 5 étoiles5/5 (1)
- Harmonisation Energétique des Lieux: Habitat et haut-lieux sacrés 2020D'EverandHarmonisation Energétique des Lieux: Habitat et haut-lieux sacrés 2020Évaluation : 2.5 sur 5 étoiles2.5/5 (3)
- Comment perdre la graisse du ventre En français/ How To Lose Belly Fat In FrenchD'EverandComment perdre la graisse du ventre En français/ How To Lose Belly Fat In FrenchPas encore d'évaluation
- Régime Cétogène: Le Guide Ultime Pour Maîtriser Une Perte De Poids Saine (Recettes De Régime Céto)D'EverandRégime Cétogène: Le Guide Ultime Pour Maîtriser Une Perte De Poids Saine (Recettes De Régime Céto)Pas encore d'évaluation
- La maîtrise de soi-même par l'autosuggestion consciente: la méthode CouéD'EverandLa maîtrise de soi-même par l'autosuggestion consciente: la méthode CouéPas encore d'évaluation
- Ces aliments qui font maigrir: Surveiller son alimentation pour perdre du poidsD'EverandCes aliments qui font maigrir: Surveiller son alimentation pour perdre du poidsPas encore d'évaluation
- Pour Maigrir: 67 recettes faciles de Jus de Fruits et de Légumes Crus Détox pour perdre du poidsD'EverandPour Maigrir: 67 recettes faciles de Jus de Fruits et de Légumes Crus Détox pour perdre du poidsÉvaluation : 5 sur 5 étoiles5/5 (2)
- Histoire de la psychologie scientifique: De la naissance de la psychologie à la neuropsychologie et aux champs d'application les plus actuelsD'EverandHistoire de la psychologie scientifique: De la naissance de la psychologie à la neuropsychologie et aux champs d'application les plus actuelsPas encore d'évaluation
- La réflexologie et ses bienfaits: Une médecine alternative pour avoir une bonne hygiène de vieD'EverandLa réflexologie et ses bienfaits: Une médecine alternative pour avoir une bonne hygiène de viePas encore d'évaluation
- Pendule et magnétisme: 2 guides en 1D'EverandPendule et magnétisme: 2 guides en 1Évaluation : 5 sur 5 étoiles5/5 (1)
- Pour être bien dans mon corps: Accepter le vieillissementD'EverandPour être bien dans mon corps: Accepter le vieillissementPas encore d'évaluation
- Chirologie radiesthésique: Analyse de la main et de son auraD'EverandChirologie radiesthésique: Analyse de la main et de son auraPas encore d'évaluation
- 100 Santé Smoothie Recettes Pour Détoxifier Et Pour Plus De Vitalité: (Guide Diététique Pour Perdre Du Poids Etse Sentir Bien Dans Son Corps)D'Everand100 Santé Smoothie Recettes Pour Détoxifier Et Pour Plus De Vitalité: (Guide Diététique Pour Perdre Du Poids Etse Sentir Bien Dans Son Corps)Pas encore d'évaluation
- Le TDAH chez l’adulte : Comment reconnaître & faire face au TDAH chez l’adulte en 30 étapes faciles.D'EverandLe TDAH chez l’adulte : Comment reconnaître & faire face au TDAH chez l’adulte en 30 étapes faciles.Pas encore d'évaluation
- Cahier de jeux de stimulation cognitive: Sujets Alzheimer, désorientés, démences, amnésiesD'EverandCahier de jeux de stimulation cognitive: Sujets Alzheimer, désorientés, démences, amnésiesPas encore d'évaluation
- Victoire totale sur toutes les maladiesD'EverandVictoire totale sur toutes les maladiesÉvaluation : 5 sur 5 étoiles5/5 (1)