Vous aimerez peut-être aussi
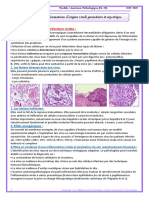
- Methode de Diagnostic 10Document55 pagesMethode de Diagnostic 10MACON824100% (1)
- M4-MB22. Virus À ARNDocument7 pagesM4-MB22. Virus À ARNسفير نعمونPas encore d'évaluation
- 3-Le Monde Microbien-ConvertiDocument40 pages3-Le Monde Microbien-ConvertiRayhane Ben SaadaPas encore d'évaluation
- Bacteriologie Systématique PDFDocument94 pagesBacteriologie Systématique PDFSofiane Lightman100% (1)
- Strategies Diag, Ostiques en VirologieDocument28 pagesStrategies Diag, Ostiques en VirologieTaoufik Ben HoumichPas encore d'évaluation
- Chapitre13 Partie2Document8 pagesChapitre13 Partie2expedit dakoPas encore d'évaluation
- Cours 4Document4 pagesCours 4mathilde.thevenin1Pas encore d'évaluation
- Cours DiagnostiqueDocument3 pagesCours Diagnostiquechekaiem monaPas encore d'évaluation
- Diagnostic. Virologique. LAFDocument52 pagesDiagnostic. Virologique. LAFmelchiPas encore d'évaluation
- Méthodes-De-Diagnostic-VirologiqueDocument4 pagesMéthodes-De-Diagnostic-VirologiquesarahPas encore d'évaluation
- Diapo PR Chaher PDFDocument51 pagesDiapo PR Chaher PDFHad EsPas encore d'évaluation
- Le Cycle EpidemiologiqueDocument43 pagesLe Cycle Epidemiologiquealibenhamida5725Pas encore d'évaluation
- Infectiologie Cours 28 Les Virus OncogenesDocument18 pagesInfectiologie Cours 28 Les Virus OncogenesPellePas encore d'évaluation
- Ecb Pus Et Liquides D Epanchement 2022Document55 pagesEcb Pus Et Liquides D Epanchement 2022IMANE EL-HAJRAOUI67% (3)
- VirologieDocument19 pagesVirologieOuld Hamadi Mohamed AliPas encore d'évaluation
- Abir ViroDocument6 pagesAbir ViroAbir LatréchePas encore d'évaluation
- M4-MB16. Diagnostic Bacteriologique (Poly)Document6 pagesM4-MB16. Diagnostic Bacteriologique (Poly)سفير نعمون100% (1)
- I - Introduction: I - 1 DéfinitionDocument13 pagesI - Introduction: I - 1 Définitionfatma aissaoui100% (1)
- Virologie Version FinaleDocument7 pagesVirologie Version FinaleOuld Hamadi Mohamed AliPas encore d'évaluation
- TP Diagnosticul VirologicDocument43 pagesTP Diagnosticul VirologicadeamaPas encore d'évaluation
- Diagnostic VirologiqueDocument5 pagesDiagnostic VirologiqueLylia Bessire100% (2)
- L'Infection Virale: E. Noel Maladies Infectieuses MicrobiologieDocument26 pagesL'Infection Virale: E. Noel Maladies Infectieuses Microbiologiejuste ben johnsonPas encore d'évaluation
- Gastroentérite ViraleDocument5 pagesGastroentérite ViraleNabih BoutaminaPas encore d'évaluation
- Chapitre 1 Virologie PDFDocument8 pagesChapitre 1 Virologie PDFazizamir charaallahPas encore d'évaluation
- Microbiologie médicale I: agents pathogènes et microbiome humainD'EverandMicrobiologie médicale I: agents pathogènes et microbiome humainÉvaluation : 5 sur 5 étoiles5/5 (1)
- CoursDocument7 pagesCourschamo chaimaPas encore d'évaluation
- Chapitre VII - L'INFLAMMATION VIRALEDocument52 pagesChapitre VII - L'INFLAMMATION VIRALEalmnaouarPas encore d'évaluation
- Support - Cours - Traitement Des Eaux 1 - Partie Microbiologique - GP3 - 2020Document12 pagesSupport - Cours - Traitement Des Eaux 1 - Partie Microbiologique - GP3 - 2020ndassisamoradanellePas encore d'évaluation
- MicrobiologieDocument63 pagesMicrobiologiealiadel100% (3)
- 3.09.11 NonhumanDocument6 pages3.09.11 NonhumanKìkøu KìkãmPas encore d'évaluation
- infections virales et etude des antivirauxDocument9 pagesinfections virales et etude des antivirauxassia bilimPas encore d'évaluation
- VIRO 2 Multiplication ViraleDocument9 pagesVIRO 2 Multiplication ViralefsxnsstoaehkbhlzygPas encore d'évaluation
- Bacterio LogieDocument37 pagesBacterio LogieMiMi NaninePas encore d'évaluation
- Cours Immunite Antivirale M1 BPMDocument8 pagesCours Immunite Antivirale M1 BPMNesrine SenouciPas encore d'évaluation
- Cours Herpes 2020-2021Document89 pagesCours Herpes 2020-2021Bouazzi IslemmPas encore d'évaluation
- 11 - Inflammation Virale, Parasitaire Et MycotiqueDocument3 pages11 - Inflammation Virale, Parasitaire Et MycotiqueNesrine DibPas encore d'évaluation
- Cytomégalovirus Et EbvDocument29 pagesCytomégalovirus Et EbvSandrine WadePas encore d'évaluation
- BactériologieDocument53 pagesBactériologieAnonymous SVP382mPas encore d'évaluation
- Le DiagnosticDocument48 pagesLe DiagnosticMerya JijiPas encore d'évaluation
- Virus de La RubéoleDocument7 pagesVirus de La RubéoleNabih BoutaminaPas encore d'évaluation
- InfectionDocument5 pagesInfectionChah Ra ZedPas encore d'évaluation
- Ouvrir UAA5 SI 5e Année 2Document48 pagesOuvrir UAA5 SI 5e Année 2b8zs895hsrPas encore d'évaluation
- Les VirusDocument20 pagesLes VirusAlassane SallPas encore d'évaluation
- Le Thème:: Séro - Diagnostic CMVDocument19 pagesLe Thème:: Séro - Diagnostic CMVNawal ArbiPas encore d'évaluation
- Virus CytomegalovirusDocument15 pagesVirus Cytomegalovirusamos kagonePas encore d'évaluation
- Hemo CultureDocument19 pagesHemo CulturesafemindPas encore d'évaluation
- Varicelle SomipevDocument48 pagesVaricelle SomipevGoGoPenguinVALPas encore d'évaluation
- Les OpportunistesDocument8 pagesLes OpportunistesShaïma BMAPas encore d'évaluation
- Le Dg. Virologique T.P. 2017 Complet 1Document56 pagesLe Dg. Virologique T.P. 2017 Complet 1adeamaPas encore d'évaluation
- 3 DG Infections ViralesDocument46 pages3 DG Infections ViralesAmenElleh ShilPas encore d'évaluation
- Diagnostic BacteriologiqueDocument68 pagesDiagnostic Bacteriologiqueabdellah abdelkhalek100% (1)
- bs-s4Document19 pagesbs-s4hasinasouyadPas encore d'évaluation
- Diagnostic IndirectDocument9 pagesDiagnostic IndirectemmaPas encore d'évaluation
- Cours Virologie M1Document59 pagesCours Virologie M1Koguep TonnyPas encore d'évaluation
- 01 Introduction A L'immunologie PDFDocument8 pages01 Introduction A L'immunologie PDFNabyl BektachePas encore d'évaluation
- Ecb Des PusDocument41 pagesEcb Des PusCharly MitanoPas encore d'évaluation
- Microbiologie, Immunologie Et Hygià Ne 22-23Document146 pagesMicrobiologie, Immunologie Et Hygià Ne 22-23Ranya ChayatPas encore d'évaluation
- InfectionDocument22 pagesInfectionحكم و نوادر إسلامية100% (1)
- Rage 1Document63 pagesRage 1bouziàne àbedPas encore d'évaluation
- ChlamydiaDocument35 pagesChlamydiaIkram El AhmerPas encore d'évaluation
- AntigenesDocument6 pagesAntigenesfsxnsstoaehkbhlzygPas encore d'évaluation
- CytokinesDocument6 pagesCytokinesfsxnsstoaehkbhlzygPas encore d'évaluation
- Chapitre 3 Physiologie BacterienneDocument9 pagesChapitre 3 Physiologie BacteriennefsxnsstoaehkbhlzygPas encore d'évaluation
- Cours 1 Introduction À BIDocument32 pagesCours 1 Introduction À BIfsxnsstoaehkbhlzygPas encore d'évaluation
- Cours 7 OMGDocument27 pagesCours 7 OMGfsxnsstoaehkbhlzygPas encore d'évaluation
- Cours 4 Introduction À La Génomique 2021Document10 pagesCours 4 Introduction À La Génomique 2021fsxnsstoaehkbhlzygPas encore d'évaluation
- Cours 2 Champs D'application Et Données BiologiquesDocument25 pagesCours 2 Champs D'application Et Données BiologiquesfsxnsstoaehkbhlzygPas encore d'évaluation
- Chapitre 1 Et 2Document27 pagesChapitre 1 Et 2vernetemma02Pas encore d'évaluation
- PolyTPBR2023 - 2024 BPRDocument9 pagesPolyTPBR2023 - 2024 BPRmahassen meskiniPas encore d'évaluation
- Epreuve de Diagnostic Serologique 1 - 051413Document3 pagesEpreuve de Diagnostic Serologique 1 - 051413Dan NdiPas encore d'évaluation
- Les Contrôles Nécessaires en Immunohistochimie: de La Recherche Au DiagnosticDocument11 pagesLes Contrôles Nécessaires en Immunohistochimie: de La Recherche Au Diagnostichaemophilus10Pas encore d'évaluation
- TP BiomolDocument6 pagesTP Biomolkamdemliemou007Pas encore d'évaluation
- TD reaction Ag-Ac. immunochimieDocument89 pagesTD reaction Ag-Ac. immunochimiehamzaabzou154Pas encore d'évaluation
- TP de Génétique TP B11. Outils Pour L'étude Des Génomes: I. Etablir Une Carte de RestrictionDocument10 pagesTP de Génétique TP B11. Outils Pour L'étude Des Génomes: I. Etablir Une Carte de Restrictionmoussodjirebecca9Pas encore d'évaluation