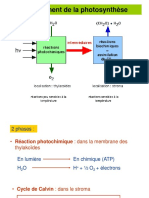
Vous aimerez peut-être aussi
- Plante c4, CamDocument22 pagesPlante c4, CamJomana Jihed100% (3)
- Bac 2023 - SVT - Corrigé - Jour 2Document4 pagesBac 2023 - SVT - Corrigé - Jour 2LETUDIANT100% (3)
- Metabolisme Bacterien Et FongiqueDocument5 pagesMetabolisme Bacterien Et FongiqueaspectmPas encore d'évaluation
- Photosynthèse ELBHAR IBRAHIMDocument60 pagesPhotosynthèse ELBHAR IBRAHIMzah ahmPas encore d'évaluation
- Kola FaneDocument9 pagesKola FaneSoumanaPas encore d'évaluation
- Photo RespirationDocument16 pagesPhoto RespirationMed OutouliPas encore d'évaluation
- CHAP1 - PV - Chap2 - Nutrition Carbonée Part2Document7 pagesCHAP1 - PV - Chap2 - Nutrition Carbonée Part2Sagacious IvejutenPas encore d'évaluation
- Relation InterspécifiqueDocument65 pagesRelation InterspécifiqueSabrina BrighetPas encore d'évaluation
- CHAP1 - PV - Chap2 - Nutrition Carbonée Part1Document10 pagesCHAP1 - PV - Chap2 - Nutrition Carbonée Part1Sagacious Ivejuten100% (1)
- Exposi Plantes CamDocument13 pagesExposi Plantes CamRahima BelmelPas encore d'évaluation
- Nutrition Carbonée Et Approvisionnement Energétique - CompletDocument31 pagesNutrition Carbonée Et Approvisionnement Energétique - CompletYassmine AbramPas encore d'évaluation
- BB 2 PhotosyncoDocument2 pagesBB 2 Photosyncohadj brahimPas encore d'évaluation
- PhotosyntheseDocument40 pagesPhotosynthesestingx50% (2)
- 2002 Nov Dec 258 259 NT9 DesbrieresDocument6 pages2002 Nov Dec 258 259 NT9 DesbrieresKhaled IdouPas encore d'évaluation
- l12 Microbio Chap5Document4 pagesl12 Microbio Chap5Alhassane BahPas encore d'évaluation
- ChloroplasteDocument4 pagesChloroplasteHugo PerazziPas encore d'évaluation
- Chapitre II Partie 2Document14 pagesChapitre II Partie 2KahinaPas encore d'évaluation
- Photorespiration - Définition Illustrée Et Explications1Document3 pagesPhotorespiration - Définition Illustrée Et Explications1أيوب زكريPas encore d'évaluation
- Les Stratégies de Photosynthèse: Les Plantes C3 - C4 Et CamDocument12 pagesLes Stratégies de Photosynthèse: Les Plantes C3 - C4 Et CamEl Mehdi El BaharPas encore d'évaluation
- CH5.Photosynthèse 2017Document26 pagesCH5.Photosynthèse 2017khammar hichemPas encore d'évaluation
- Photosynthese PascalDocument55 pagesPhotosynthese Pascalmeno donPas encore d'évaluation
- Chapitre 2: Croissance Bactérienne: A-Matériel UtiliséDocument8 pagesChapitre 2: Croissance Bactérienne: A-Matériel UtiliséKamel BourasPas encore d'évaluation
- La Photosynthèse Cours de Biochimie 2eme Année Pharmacie DR CHERIETDocument15 pagesLa Photosynthèse Cours de Biochimie 2eme Année Pharmacie DR CHERIETsarahsora55Pas encore d'évaluation
- Plante c4 PDFDocument32 pagesPlante c4 PDFقادة قناية100% (4)
- Chap1 PhotosyntheseDocument2 pagesChap1 PhotosyntheseAbdoulaye Junior DialloPas encore d'évaluation
- Spe Sciences Vie Terre CorrigeDocument6 pagesSpe Sciences Vie Terre Corrigek9mystakesPas encore d'évaluation
- La Production Microbienne Des PolysaccharidesDocument16 pagesLa Production Microbienne Des PolysaccharidesAymen BoudiafPas encore d'évaluation
- Chp2 ABDocument8 pagesChp2 ABwassiouadjayi8Pas encore d'évaluation
- Bacterio - UEBIO238 - Moodle - 1Document11 pagesBacterio - UEBIO238 - Moodle - 1LARIPas encore d'évaluation
- MetabolismeDocument34 pagesMetabolismelilyrosejegoPas encore d'évaluation
- TD3 Nutrition Metabolisme CroissanceDocument28 pagesTD3 Nutrition Metabolisme CroissanceTemtem HmPas encore d'évaluation
- Biologie Organites CellulaireDocument102 pagesBiologie Organites CellulairenouhabettachaPas encore d'évaluation
- 2 - La Cellule Végétale - FicheDocument16 pages2 - La Cellule Végétale - FicheLisa FoucherPas encore d'évaluation
- 30.09.2015 Catabolisme CellulaireDocument23 pages30.09.2015 Catabolisme CellulaireEaPas encore d'évaluation
- TD2 PHYSIOLOGIE VEGETALE - Correction DORGELESDocument2 pagesTD2 PHYSIOLOGIE VEGETALE - Correction DORGELESSahouin Claude Medard0% (1)
- Valorisation Des Espèces MicroalguesDocument6 pagesValorisation Des Espèces Microalguesf.ezzahraeadnaniPas encore d'évaluation
- Resume Sciences Bilogique Pour L'ingenieurDocument37 pagesResume Sciences Bilogique Pour L'ingenieurSara AdPas encore d'évaluation
- Biocellulaire 2 Partie-1Document29 pagesBiocellulaire 2 Partie-1Jdiid Raî ABDELKRIMPas encore d'évaluation
- 5 La Membrane PlasmiqueDocument11 pages5 La Membrane PlasmiqueKí KŷuśặkuPas encore d'évaluation
- Bio Cell (Semaine 4)Document77 pagesBio Cell (Semaine 4)Maryem BouzebbarPas encore d'évaluation
- S217 Chapitre 6 Les MosissuresDocument64 pagesS217 Chapitre 6 Les MosissuresBadr GourmahPas encore d'évaluation
- Cours de Botanique 2022Document103 pagesCours de Botanique 2022Hikari NantePas encore d'évaluation
- 2 Bac FR PC (1° Semestre)Document96 pages2 Bac FR PC (1° Semestre)mohamedPas encore d'évaluation
- Algues BrunesDocument41 pagesAlgues BruneshouriaPas encore d'évaluation
- Liberation de L Energie Emmagasinee Dans La Matiere Organique Cours 3Document18 pagesLiberation de L Energie Emmagasinee Dans La Matiere Organique Cours 3Mohamed WidadPas encore d'évaluation
- Production de La Matière Organique Et Flux D'énergieDocument22 pagesProduction de La Matière Organique Et Flux D'énergieViperDi AB100% (1)
- Colonne Wino 11-22 1Document67 pagesColonne Wino 11-22 1daniel kambaPas encore d'évaluation
- Les Algues Ou Les Phycophytes: I. Caractéristiques Cytologiques Des AlguesDocument27 pagesLes Algues Ou Les Phycophytes: I. Caractéristiques Cytologiques Des AlguesLaura West ManierPas encore d'évaluation
- ChloroplastDocument4 pagesChloroplastZahraPas encore d'évaluation
- Rapport de StageDocument16 pagesRapport de Stagepraetf100% (1)
- La PHOTOSYNTHÈSE PAR DES CHLOROPLASTES ISOLÉSDocument3 pagesLa PHOTOSYNTHÈSE PAR DES CHLOROPLASTES ISOLÉSElia FadelPas encore d'évaluation
- 3 Les AlcaloidesDocument11 pages3 Les AlcaloidesInes DaoudiPas encore d'évaluation
- UE2 - Biologie Cellulaire - Production D - Énergie Dans La CelluleDocument16 pagesUE2 - Biologie Cellulaire - Production D - Énergie Dans La Cellulesun-nee-chan9Pas encore d'évaluation
- Chapitre 3 Les AlguesDocument26 pagesChapitre 3 Les AlguesLaura West ManierPas encore d'évaluation
- Photosynthese 2 S3 Licence HorticultureDocument43 pagesPhotosynthese 2 S3 Licence HorticultureHicham Ait BouhouPas encore d'évaluation
- Consommation de La Matière Organique Et Flux D'énergie - Cour1Document13 pagesConsommation de La Matière Organique Et Flux D'énergie - Cour1timmary66Pas encore d'évaluation
- Terpènes Et Biosynthèse TerpéniqueDocument15 pagesTerpènes Et Biosynthèse TerpéniqueRẵmîRandyHassad100% (1)
- Chap 9 Et 10Document11 pagesChap 9 Et 10ĄbdÖù CatalonPas encore d'évaluation
- Membranes cellulaires: Les Grands Articles d'UniversalisD'EverandMembranes cellulaires: Les Grands Articles d'UniversalisPas encore d'évaluation
- Ilford Fp4 PlusDocument6 pagesIlford Fp4 PlusMatthieu BEGELPas encore d'évaluation
- (Géopolitique Mondiale) Wenu Becker - Diplomatie Et Sécurité en RDC de 1960 À 1965-L'Harmattan (2017)Document141 pages(Géopolitique Mondiale) Wenu Becker - Diplomatie Et Sécurité en RDC de 1960 À 1965-L'Harmattan (2017)Matthieu MatiyaboPas encore d'évaluation
- Energies RenouvelablesDocument16 pagesEnergies RenouvelablesMohamed WarkzizPas encore d'évaluation
- Jeux D'ombresDocument388 pagesJeux D'ombresArnaud BallPas encore d'évaluation
- Oral Français LL4 - Vénus AnadyomèneDocument6 pagesOral Français LL4 - Vénus Anadyomènejulie khavathPas encore d'évaluation
- Microbiologie PrédictiveDocument9 pagesMicrobiologie Prédictivevague20000% (1)
- Soutenance Projet EI5 - Ruche IntelligenteDocument34 pagesSoutenance Projet EI5 - Ruche IntelligenteTMauginPas encore d'évaluation
- Correction TD Alzheimer (Début)Document4 pagesCorrection TD Alzheimer (Début)kettouchePas encore d'évaluation
- Devoir de Contrôle N°3 - SVT - 2ème SC (2006-2007) Mme Habli RajaDocument3 pagesDevoir de Contrôle N°3 - SVT - 2ème SC (2006-2007) Mme Habli RajaChihab GhlalaPas encore d'évaluation
- TP ExtractionDocument6 pagesTP ExtractionInes DaoudiPas encore d'évaluation
- Busch Instruction Manual KB KC 0040 D FR 0870152114 PDFDocument22 pagesBusch Instruction Manual KB KC 0040 D FR 0870152114 PDFaziz youssfiPas encore d'évaluation
- EVCP OphtalmoDocument31 pagesEVCP OphtalmoBouchoucha SakrPas encore d'évaluation
- Epreuv Bac A 2012Document1 pageEpreuv Bac A 2012FOGNO FOTSOPas encore d'évaluation
- Gestion de StockDocument6 pagesGestion de Stockexpert diag autoPas encore d'évaluation
- Mieux Comprendre LDocument5 pagesMieux Comprendre LAchraf CpnPas encore d'évaluation
- Franc 5kl Ch1 Vadushina Rus Bel 2020Document139 pagesFranc 5kl Ch1 Vadushina Rus Bel 2020Мария МисюкPas encore d'évaluation
- P1AR00 - Captain Jacks Restaurant PiratesDocument28 pagesP1AR00 - Captain Jacks Restaurant PiratesBruce TandirePas encore d'évaluation
- 4e Classe, Partie A, Questions de RévisionDocument335 pages4e Classe, Partie A, Questions de RévisionScribdTranslationsPas encore d'évaluation
- 3 CCTP HEM BatimentDocument197 pages3 CCTP HEM BatimentGosson Yenipoho Aboubacar SoroPas encore d'évaluation
- Serveur WES Installation V2Document14 pagesServeur WES Installation V2Johann LelandaisPas encore d'évaluation
- Cas CliniqueDocument3 pagesCas Cliniquesamyktg44Pas encore d'évaluation
- La Médecine Des Pharaons. Magie Et Science Médicale Dans L'égypte AncienneDocument27 pagesLa Médecine Des Pharaons. Magie Et Science Médicale Dans L'égypte AncienneGuillermo Benitez100% (3)
- Bac 2014 Bac Tech Epreuve Physique Session Controle Correction PDFDocument5 pagesBac 2014 Bac Tech Epreuve Physique Session Controle Correction PDFmomme deyahiPas encore d'évaluation
- Plan de Visite 2018Document2 pagesPlan de Visite 2018Benoît AgassantPas encore d'évaluation
- Poétique D'aristote (Nouv Éd Rev (... ) Aristote (0384-0322 bpt6k70607gDocument57 pagesPoétique D'aristote (Nouv Éd Rev (... ) Aristote (0384-0322 bpt6k70607gCapela da FilôPas encore d'évaluation
- Embaucher Un Salarie en Elevage VF Maj2016Document16 pagesEmbaucher Un Salarie en Elevage VF Maj2016gyressestellaPas encore d'évaluation
- Studio Mumbai, Bijoy & Priya Jain, ArchitectesDocument12 pagesStudio Mumbai, Bijoy & Priya Jain, Architectesben_65Pas encore d'évaluation
- Liste Fournitures Scolaires CE1 2023 2024 1Document3 pagesListe Fournitures Scolaires CE1 2023 2024 1diagonale ecolePas encore d'évaluation
- La Technique "S'efface"Document20 pagesLa Technique "S'efface"TRIBYPas encore d'évaluation