Vous aimerez peut-être aussi
- Photosynthèse: Les Grands Articles d'UniversalisD'EverandPhotosynthèse: Les Grands Articles d'UniversalisPas encore d'évaluation
- Bio Cell - Part 4Document20 pagesBio Cell - Part 4Zineb hmt27Pas encore d'évaluation
- 2 - La Cellule Végétale - FicheDocument16 pages2 - La Cellule Végétale - FicheLisa FoucherPas encore d'évaluation
- Membranes cellulaires: Les Grands Articles d'UniversalisD'EverandMembranes cellulaires: Les Grands Articles d'UniversalisPas encore d'évaluation
- La Cellule Végétale - CHEIKHDocument56 pagesLa Cellule Végétale - CHEIKHbyaya7431Pas encore d'évaluation
- La Photosynthèse Cours de Biochimie 2eme Année Pharmacie DR CHERIETDocument15 pagesLa Photosynthèse Cours de Biochimie 2eme Année Pharmacie DR CHERIETsarahsora55Pas encore d'évaluation
- CHAP1 - PV - Chap2 - Nutrition Carbonée Part1Document10 pagesCHAP1 - PV - Chap2 - Nutrition Carbonée Part1Sagacious Ivejuten100% (1)
- Biologie CellulaireDocument23 pagesBiologie CellulaireKévin Colin100% (3)
- 3.1 Rappels Sur Les Types Trophiques: 3) La PhotosynthèseDocument23 pages3.1 Rappels Sur Les Types Trophiques: 3) La PhotosynthèseAmanda LaihongtingPas encore d'évaluation
- Chapitre 2 BVDocument10 pagesChapitre 2 BVThe AngelPas encore d'évaluation
- La PhotosynthèseDocument2 pagesLa PhotosynthèseR 23Pas encore d'évaluation
- Diversité Et Complémentarité Des MétabolismesDocument10 pagesDiversité Et Complémentarité Des MétabolismesJosé Ahanda NguiniPas encore d'évaluation
- Identite BiologiaueDocument15 pagesIdentite BiologiauePessu EricPas encore d'évaluation
- Physiologie Végétale L2 Fonkou 2023Document48 pagesPhysiologie Végétale L2 Fonkou 2023Leonel RochePas encore d'évaluation
- La Cellule EucaryoteDocument9 pagesLa Cellule Eucaryoteherrere64Pas encore d'évaluation
- 2de T1 C2 BilanDocument7 pages2de T1 C2 BilanDupontPas encore d'évaluation
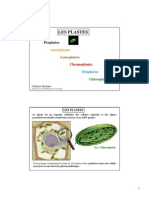
- 6 Les Plastes CaffarriDocument14 pages6 Les Plastes Caffarriرمضان كريمPas encore d'évaluation
- La Cellule Algale A Des Caractéristiques RemarquablesDocument1 pageLa Cellule Algale A Des Caractéristiques RemarquablesAhmedPas encore d'évaluation
- Biochimie AlimentaireDocument63 pagesBiochimie AlimentaireCiobanu Vladislav100% (1)
- Chapitre 1 - Suite Cellules Eucaryote Poly ElèvesDocument8 pagesChapitre 1 - Suite Cellules Eucaryote Poly Elèvesmaximedijoux6Pas encore d'évaluation
- Physiologie Vegetale 2 S4Document20 pagesPhysiologie Vegetale 2 S4hassan aitPas encore d'évaluation
- Biologie La CelluleDocument12 pagesBiologie La Cellule6z8nbjv76vPas encore d'évaluation
- Saccharose Ou AmidonDocument4 pagesSaccharose Ou AmidonKhadija El aroussiPas encore d'évaluation
- Cytoplasm eDocument15 pagesCytoplasm eyasPas encore d'évaluation
- Production D'energie PDFDocument10 pagesProduction D'energie PDFSoubere MahamoudPas encore d'évaluation
- Cours Biologie Végétale Chap 1 2 3Document37 pagesCours Biologie Végétale Chap 1 2 3Nabil holmes100% (6)
- Chapitre 3 Les AlguesDocument26 pagesChapitre 3 Les AlguesLaura West ManierPas encore d'évaluation
- Cytoplasme Bio Cell Sbaa 2024Document25 pagesCytoplasme Bio Cell Sbaa 2024habessongPas encore d'évaluation
- ExtraitDocument8 pagesExtraitAshraf Ouro-adohiPas encore d'évaluation
- UE2 - Biologie Cellulaire - Production D - Énergie Dans La CelluleDocument16 pagesUE2 - Biologie Cellulaire - Production D - Énergie Dans La Cellulesun-nee-chan9Pas encore d'évaluation
- 9 CLASSE DE Tle C-1Document103 pages9 CLASSE DE Tle C-1Njiasse MaazouPas encore d'évaluation
- 1.cours Biologie CellulaireDocument63 pages1.cours Biologie Cellulaireبدر الذهابيPas encore d'évaluation
- Les Mitochondries Et Les ChloroplastesDocument20 pagesLes Mitochondries Et Les ChloroplastesMbaye AbouPas encore d'évaluation
- Bio Cell (Semaine 4)Document77 pagesBio Cell (Semaine 4)Maryem BouzebbarPas encore d'évaluation
- Chapitre I Structure de La CelluleDocument107 pagesChapitre I Structure de La CelluleDescartes KpannePas encore d'évaluation
- Plaste PDFDocument23 pagesPlaste PDFrashn7100% (2)
- La CelluleDocument26 pagesLa CellulesaadiaPas encore d'évaluation
- 30.09.2015 Catabolisme CellulaireDocument23 pages30.09.2015 Catabolisme CellulaireEaPas encore d'évaluation
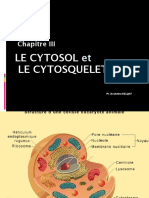
- CH3 - Cytosol-Cytosquelette - B.BDocument47 pagesCH3 - Cytosol-Cytosquelette - B.Bone asslanPas encore d'évaluation
- Mito WRDocument3 pagesMito WRAbir M'ghariPas encore d'évaluation
- Les CellulesDocument5 pagesLes CellulesyoussefPas encore d'évaluation
- TD7 Les Organites de La Cellule PDFDocument42 pagesTD7 Les Organites de La Cellule PDFZineb DahmaniPas encore d'évaluation
- Cours No 03-La Cellule (Suite)Document8 pagesCours No 03-La Cellule (Suite)Feriel Feriel0% (1)
- Chapitre X Les MitochondriesDocument12 pagesChapitre X Les MitochondriesMichael Alafakis100% (1)
- Cours No 02-La CelluleDocument13 pagesCours No 02-La CelluleFeriel FerielPas encore d'évaluation
- Membrane PlasmiqueDocument7 pagesMembrane PlasmiqueSara BoudjemaiPas encore d'évaluation
- Cours BCG UFR2SDocument19 pagesCours BCG UFR2SSirandou DiaoPas encore d'évaluation
- 9 MitochondriesDocument42 pages9 MitochondriesTyl HyuPas encore d'évaluation
- Chap 2 MPDocument7 pagesChap 2 MPdramohandlhadjPas encore d'évaluation
- La Mitochondrie Est Un Organite Clos Possédant LesDocument32 pagesLa Mitochondrie Est Un Organite Clos Possédant LesFaresPas encore d'évaluation
- Cellue 2022 SamassekouDocument13 pagesCellue 2022 SamassekouMohamed DickoPas encore d'évaluation
- Chapitre1 Biochimie Cellulaire Et FonctionnelleDocument12 pagesChapitre1 Biochimie Cellulaire Et Fonctionnelledraboissouf174Pas encore d'évaluation
- REF Cytologie PVDocument11 pagesREF Cytologie PVHelena HINTZER--FALCOPas encore d'évaluation
- Exposé de Cytologie: ThèmeDocument41 pagesExposé de Cytologie: ThèmeFaresPas encore d'évaluation
- Chap. 3. LES ORGANITES CELLULAIRESDocument18 pagesChap. 3. LES ORGANITES CELLULAIRESbanassimconcepta6Pas encore d'évaluation
- Noyau (Biologie) - WikipédiaDocument22 pagesNoyau (Biologie) - Wikipédia4xxstan2022Pas encore d'évaluation
- Partie IDocument3 pagesPartie IhmzPas encore d'évaluation
- La Cellule EucaryoteDocument2 pagesLa Cellule EucaryoteIngrid Marchaux100% (1)
- ChloroplasteDocument4 pagesChloroplasteHugo PerazziPas encore d'évaluation
- 30 Astuces Pour Gagner Du Temps Avec ExcelDocument11 pages30 Astuces Pour Gagner Du Temps Avec ExceldjennatialiPas encore d'évaluation
- Génie GénitiqueDocument3 pagesGénie GénitiqueJdiid Raî ABDELKRIM100% (1)
- Astronom I eDocument3 pagesAstronom I eJdiid Raî ABDELKRIMPas encore d'évaluation
- MondialisationDocument9 pagesMondialisationJdiid Raî ABDELKRIMPas encore d'évaluation
- MondialisationDocument9 pagesMondialisationJdiid Raî ABDELKRIMPas encore d'évaluation
- SVI Correspondance Modules14 15Document2 pagesSVI Correspondance Modules14 15Don YaPas encore d'évaluation
- Archéologie de La Violence - Pierre ClastresDocument27 pagesArchéologie de La Violence - Pierre ClastresleefalkPas encore d'évaluation
- A Case Study of Ground Improvement Using PiersDocument5 pagesA Case Study of Ground Improvement Using PiersAshraf Gomah Mohamed ElshamandyPas encore d'évaluation
- Laplace - Oeuvres Complètes Tome IIDocument418 pagesLaplace - Oeuvres Complètes Tome IIOmar KassemPas encore d'évaluation
- Intro+aperçu Géographique OuenzaDocument2 pagesIntro+aperçu Géographique OuenzaSõlã KhedPas encore d'évaluation
- ArgiiileDocument47 pagesArgiiileraoufPas encore d'évaluation
- Premiers Humains, Premières Migrations: Histoire Les Débuts de L'humanité Fiche 1Document2 pagesPremiers Humains, Premières Migrations: Histoire Les Débuts de L'humanité Fiche 1api-468372798Pas encore d'évaluation
- Astrologie Karmique PDFDocument2 pagesAstrologie Karmique PDFrastacourage67% (3)
- 669 - Martine CosmeticsDocument1 page669 - Martine CosmeticsKelly Silva CastroPas encore d'évaluation
- Guide D'apprentissageDocument31 pagesGuide D'apprentissageAdil LamPas encore d'évaluation
- Applications Des Dispositifs ET DES Methodes DE Traitement DES Mesures Electriques Dans LES Regions DE Chott EL Gharbi ET Chott CherguiDocument96 pagesApplications Des Dispositifs ET DES Methodes DE Traitement DES Mesures Electriques Dans LES Regions DE Chott EL Gharbi ET Chott CherguiIlh BrighPas encore d'évaluation
- Daufrene K Taping PDFDocument47 pagesDaufrene K Taping PDFMjidKarim100% (1)
- BELKAMEL Aissa PDFDocument126 pagesBELKAMEL Aissa PDFYoucef BenferdiPas encore d'évaluation
- Le Monde Des Teintures Naturelles Nouvelle Édition PrésentationDocument2 pagesLe Monde Des Teintures Naturelles Nouvelle Édition PrésentationØüssÂmāPas encore d'évaluation
- Comment Développer Votre Attention Et Votre Mémoire PDFDocument37 pagesComment Développer Votre Attention Et Votre Mémoire PDFFresnel FulgencePas encore d'évaluation
- Les Sources Thermales en AlgérieDocument3 pagesLes Sources Thermales en AlgérieImene Kartoby100% (1)
- Guide Bassin ArcachonDocument100 pagesGuide Bassin ArcachonChe MaPas encore d'évaluation
- 04 - Pression Des Terres - Solutionnaire PDFDocument26 pages04 - Pression Des Terres - Solutionnaire PDFmeksi aliPas encore d'évaluation
- Archidoxe Magique ParacelseDocument55 pagesArchidoxe Magique ParacelseSpartakus FreeMann100% (3)
- Les Indices BiologiqueDocument47 pagesLes Indices BiologiqueLina Ray Ben91% (11)
- Exercices Sur Lastronomie Avec Corrigé - 2 PDFDocument6 pagesExercices Sur Lastronomie Avec Corrigé - 2 PDFAnonymous 80p9OVPas encore d'évaluation
- AgendaDocument1 pageAgendafodekambalouPas encore d'évaluation
- Physique Et Réalité - Le Temps Existe-T-IlDocument35 pagesPhysique Et Réalité - Le Temps Existe-T-IlMichael64Pas encore d'évaluation
- BiophysiqueDocument5 pagesBiophysiqueJihan SafwaPas encore d'évaluation
- Quel Projet Urbain Pour Un Retour de La Ville A La Mer SkikdaDocument356 pagesQuel Projet Urbain Pour Un Retour de La Ville A La Mer SkikdaKHELIFA100% (2)
- TFCDocument92 pagesTFCGrace Emile Moelet Goma67% (3)