Vous aimerez peut-être aussi
- Reticulum Et GolgiDocument10 pagesReticulum Et Golgisatisfying541Pas encore d'évaluation
- Chapitre VI - Système Endomembranaire - Janvier 2024Document11 pagesChapitre VI - Système Endomembranaire - Janvier 2024Swayli HossamPas encore d'évaluation
- Partie 2. Cours de BioCell 1ASNVDocument17 pagesPartie 2. Cours de BioCell 1ASNVايمن معطااللهPas encore d'évaluation
- Cours de Cytologie, Embryologie Et Genetique Medicale by MBAILASSEM MBAINAIDA GEORGESDocument102 pagesCours de Cytologie, Embryologie Et Genetique Medicale by MBAILASSEM MBAINAIDA GEORGESMãhąmãţ Hãbįb Mõųşşã ĄhmąţPas encore d'évaluation
- Chapitre I Structure de La CelluleDocument107 pagesChapitre I Structure de La CelluleDescartes KpannePas encore d'évaluation
- REF Cytologie PVDocument11 pagesREF Cytologie PVHelena HINTZER--FALCOPas encore d'évaluation
- La CelluleDocument26 pagesLa CellulesaadiaPas encore d'évaluation
- Le Systeme Endomembranaire Réticulum Endoplasmique, Appareil de Golgi, LysosomesDocument22 pagesLe Systeme Endomembranaire Réticulum Endoplasmique, Appareil de Golgi, LysosomesAy ManPas encore d'évaluation
- Les Compartiments IntracellulairesDocument60 pagesLes Compartiments Intracellulairesmarie dominique patricia kouassiPas encore d'évaluation
- 9 CLASSE DE Tle C-1Document103 pages9 CLASSE DE Tle C-1Njiasse MaazouPas encore d'évaluation
- Cours BCG UFR2SDocument19 pagesCours BCG UFR2SSirandou DiaoPas encore d'évaluation
- Cytoplasm eDocument15 pagesCytoplasm eyasPas encore d'évaluation
- Cours 3. Membrane PlasmiqueDocument14 pagesCours 3. Membrane PlasmiqueNã BîlPas encore d'évaluation
- Compartiments Cellulaires Partie 1Document68 pagesCompartiments Cellulaires Partie 1anasbwedaPas encore d'évaluation
- Snv-Biologie Cellulaire-Les Membranes CellulairesDocument5 pagesSnv-Biologie Cellulaire-Les Membranes Cellulairesamira messoudPas encore d'évaluation
- Cours Système Endomembranaire 2021 2022Document9 pagesCours Système Endomembranaire 2021 2022samysaib79Pas encore d'évaluation
- La CelluleDocument6 pagesLa Celluleikram dahdouhPas encore d'évaluation
- La Cellule Végétale - CHEIKHDocument56 pagesLa Cellule Végétale - CHEIKHbyaya7431Pas encore d'évaluation
- Expose Du Groupe GDocument4 pagesExpose Du Groupe Gdorechristine04Pas encore d'évaluation
- Biochimie AlimentaireDocument63 pagesBiochimie AlimentaireCiobanu Vladislav100% (1)
- TD7 Les Organites de La Cellule PDFDocument42 pagesTD7 Les Organites de La Cellule PDFZineb DahmaniPas encore d'évaluation
- Le Réticulum Endoplasmique - 2Document45 pagesLe Réticulum Endoplasmique - 2ilyes.jilouPas encore d'évaluation
- Cours Membrane Plasmique & Hyaloplasme Microsoft Office WordDocument19 pagesCours Membrane Plasmique & Hyaloplasme Microsoft Office WordJoan Jean Jephthé Babo100% (4)
- La Cellule Algale A Des Caractéristiques RemarquablesDocument1 pageLa Cellule Algale A Des Caractéristiques RemarquablesAhmedPas encore d'évaluation
- Structure Des Membranes Lipidiques 1Document51 pagesStructure Des Membranes Lipidiques 1Abdellah EL AnsariPas encore d'évaluation
- Chapitre 02 La Membrane CellulaireDocument13 pagesChapitre 02 La Membrane Cellulairecqkj7sn9kxPas encore d'évaluation
- Cellue 2022 SamassekouDocument13 pagesCellue 2022 SamassekouMohamed DickoPas encore d'évaluation
- BIO 111 - CHAP - VI - VII - Et - VIIIDocument9 pagesBIO 111 - CHAP - VI - VII - Et - VIIIBerthe Aristide Ngue NkongoPas encore d'évaluation
- La Cellule EucaryoteDocument9 pagesLa Cellule Eucaryoteherrere64Pas encore d'évaluation
- Chapitre Ii Etude de La Cellule Eucaryote Et Eucaryote PDFDocument10 pagesChapitre Ii Etude de La Cellule Eucaryote Et Eucaryote PDFقادة قناية100% (1)
- Biologie CellulaireDocument26 pagesBiologie CellulaireBourobou bourobou Chadrack gaelPas encore d'évaluation
- La Cellule EucaryoteDocument2 pagesLa Cellule EucaryoteIngrid Marchaux100% (1)
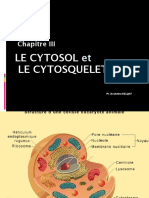
- CH3 - Cytosol-Cytosquelette - B.BDocument47 pagesCH3 - Cytosol-Cytosquelette - B.Bone asslanPas encore d'évaluation
- Bcis 3Document25 pagesBcis 3rimas23rimas14Pas encore d'évaluation
- Organisation Generale de La Cellule 2016-2017 PDFDocument3 pagesOrganisation Generale de La Cellule 2016-2017 PDFAnonymous wVr908lBcK0% (1)
- 1.cours Biologie CellulaireDocument63 pages1.cours Biologie Cellulaireبدر الذهابيPas encore d'évaluation
- Structure BactérienneDocument72 pagesStructure BactériennekhouildiPas encore d'évaluation
- BIO CEL Chap 4 - 3 Système Endomembranaire 20-21-1Document20 pagesBIO CEL Chap 4 - 3 Système Endomembranaire 20-21-1fatimaarrach59Pas encore d'évaluation
- Chapitre 1 - Suite Cellules Eucaryote Poly ElèvesDocument8 pagesChapitre 1 - Suite Cellules Eucaryote Poly Elèvesmaximedijoux6Pas encore d'évaluation
- Réticulum Endoplasmique OranDocument7 pagesRéticulum Endoplasmique OranHaitam El OuahabiPas encore d'évaluation
- Membrane PlasmiqueDocument8 pagesMembrane PlasmiqueGaston CarinePas encore d'évaluation
- Cours Systeme EndomembranaireDocument9 pagesCours Systeme EndomembranaireIOana Borca100% (1)
- Cellule Et TissusDocument44 pagesCellule Et TissusBANDADILAHCENPas encore d'évaluation
- Biologie Cellulaire Et MoléculaireDocument10 pagesBiologie Cellulaire Et Moléculaireaboubaker fantasyPas encore d'évaluation
- Intrebari AnatomieDocument15 pagesIntrebari AnatomieAlexandra Ana-Maria IancuPas encore d'évaluation
- Cytoplasme 4Document27 pagesCytoplasme 4Oumarou KontaPas encore d'évaluation
- Atlas en Couleur DhistologieDocument55 pagesAtlas en Couleur DhistologieDenisco NkemgnePas encore d'évaluation
- Les CellulesDocument3 pagesLes Cellulesoserobin21Pas encore d'évaluation
- 1 Anatomie Physiologie CelluleDocument30 pages1 Anatomie Physiologie CellulefatimazzahrakikiPas encore d'évaluation
- CytologieDocument43 pagesCytologieMouats Badreddine100% (2)
- 2021 - Cours La Cellule Version ComplèteDocument10 pages2021 - Cours La Cellule Version Complèteagathe surusPas encore d'évaluation
- ExtraitDocument8 pagesExtraitAshraf Ouro-adohiPas encore d'évaluation
- 2 - La Cellule Végétale - FicheDocument16 pages2 - La Cellule Végétale - FicheLisa FoucherPas encore d'évaluation
- Quelle Est L'ultrastructure de La Cellule Eucaryote ?Document5 pagesQuelle Est L'ultrastructure de La Cellule Eucaryote ?youseftriki7Pas encore d'évaluation
- Chap1 EucaryoteDocument4 pagesChap1 Eucaryoteslimdamak100% (1)
- Chapitre I Structure de La CelluleDocument103 pagesChapitre I Structure de La CelluleMama GueyePas encore d'évaluation
- Chapitre VII Le Reticulum Endoplasmique Et Lappareil de Golgi B8Document13 pagesChapitre VII Le Reticulum Endoplasmique Et Lappareil de Golgi B8kim namjoonPas encore d'évaluation
- 02 Revetement-CellulaireDocument6 pages02 Revetement-CellulaireSagno AgnèsPas encore d'évaluation
- Membranes cellulaires: Les Grands Articles d'UniversalisD'EverandMembranes cellulaires: Les Grands Articles d'UniversalisPas encore d'évaluation
- Surrénales: Les Grands Articles d'UniversalisD'EverandSurrénales: Les Grands Articles d'UniversalisPas encore d'évaluation
- 9e CH 4 Notes NVXDocument2 pages9e CH 4 Notes NVXHei SuPas encore d'évaluation
- GP47Document1 pageGP47xgfhdu100% (1)
- La Membrane Biologique 1 PDFDocument11 pagesLa Membrane Biologique 1 PDFsouberePas encore d'évaluation
- Le Transport MembranaireDocument17 pagesLe Transport MembranaireKoukiza TalebPas encore d'évaluation
- FC - Mitochondries Et PeroxysomesDocument17 pagesFC - Mitochondries Et PeroxysomesGabriel VerspyckPas encore d'évaluation
- 4-2-Trafic Intracellulaire Des Protéines vf1-2023Document9 pages4-2-Trafic Intracellulaire Des Protéines vf1-2023Aggoun FadouaPas encore d'évaluation
- 1BAC-Unite 3 Chapitre 2 CompletDocument8 pages1BAC-Unite 3 Chapitre 2 Completyoussefichour2Pas encore d'évaluation
- Résumé GajaanaDocument45 pagesRésumé GajaanalolipopmaronierPas encore d'évaluation
- Trafic Intracellulaire Et Compartiment CellulaireDocument24 pagesTrafic Intracellulaire Et Compartiment Cellulairecoursenligne.lille186% (7)
- Chapitre 3 PHYSIOLOGIE DES GLOBULES ROUGESDocument20 pagesChapitre 3 PHYSIOLOGIE DES GLOBULES ROUGESAlexandra SparksPas encore d'évaluation
- C Proc Et EucDocument23 pagesC Proc Et EucAya youyaPas encore d'évaluation
- Lysosome BPCDocument19 pagesLysosome BPCMzn JannahPas encore d'évaluation
- Bio CellulaireDocument44 pagesBio CellulaireTouri Na100% (1)
- Biologie Cellulaire Mai 2023Document5 pagesBiologie Cellulaire Mai 2023Jean-Yves ESTINFILPas encore d'évaluation
- DR A. AouicheDocument43 pagesDR A. Aouicheamira laggounPas encore d'évaluation
- UE2 La-Membrane-PlasmiqueDocument17 pagesUE2 La-Membrane-Plasmiquesun-nee-chan9Pas encore d'évaluation
- Memoire YbDocument204 pagesMemoire Ybegsamir1075Pas encore d'évaluation
- Cours BIO-1-Définition & Origine de La VieDocument90 pagesCours BIO-1-Définition & Origine de La VieBenjamin Plee100% (1)
- Metaux Lourds SanteDocument36 pagesMetaux Lourds Santehonore belloti100% (1)
- Appareil de Golgi 2015-2016 PDFDocument4 pagesAppareil de Golgi 2015-2016 PDFAnonymous wVr908lBcK100% (3)
- Rapport de Bio Extraction d'ADNDocument11 pagesRapport de Bio Extraction d'ADNEva BermudezPas encore d'évaluation
- Cours EntierDocument33 pagesCours EntierGayaPas encore d'évaluation
- 10-48982 Métabolisme Énergétique EMC 2008Document16 pages10-48982 Métabolisme Énergétique EMC 2008koloPas encore d'évaluation
- DS Cellule 2Document2 pagesDS Cellule 2Oudad 3Pas encore d'évaluation
- Écrire Des Questions D'examen À L'épreuve de La Tricherie - Cours Rétroactions, Inspé, Univ. Grenoble AlpesDocument7 pagesÉcrire Des Questions D'examen À L'épreuve de La Tricherie - Cours Rétroactions, Inspé, Univ. Grenoble AlpesKhaouda DrissPas encore d'évaluation
- th0238066 TheseDocument231 pagesth0238066 ThesePFEPas encore d'évaluation
- Correction Questions Ouvertes Examens AntérieursDocument32 pagesCorrection Questions Ouvertes Examens AntérieursLorenz MeurrensPas encore d'évaluation
- Khelfi 2018Document13 pagesKhelfi 2018Wafaa ZagharyPas encore d'évaluation
- 2 Cours Cyto Les Transports MBNDocument21 pages2 Cours Cyto Les Transports MBNegos013Pas encore d'évaluation
- Globules RougesDocument17 pagesGlobules RougesMounia El YousfiPas encore d'évaluation