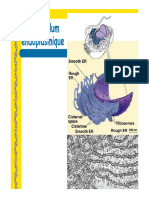
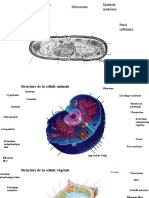
Vous aimerez peut-être aussi
- Les Lysosomes 2015-2016 PDFDocument3 pagesLes Lysosomes 2015-2016 PDFAnonymous wVr908lBcK100% (3)
- Lysosome - WikipédiaDocument13 pagesLysosome - Wikipédiagagnon.jonathan2007Pas encore d'évaluation
- Structure Des Membranes Lipidiques 1Document51 pagesStructure Des Membranes Lipidiques 1Abdellah EL AnsariPas encore d'évaluation
- Chapitre 02 La Membrane CellulaireDocument13 pagesChapitre 02 La Membrane Cellulairecqkj7sn9kxPas encore d'évaluation
- Le Systeme Endo MembranaireDocument30 pagesLe Systeme Endo MembranaireAbd El AzizPas encore d'évaluation
- 3 Membrane PlasmiqueDocument53 pages3 Membrane PlasmiqueLina RazekPas encore d'évaluation
- Chapitre 3 Maturation Des ProtéinesDocument34 pagesChapitre 3 Maturation Des Protéinesryadh2203Pas encore d'évaluation
- Réticulum Endoplasmique OranDocument7 pagesRéticulum Endoplasmique OranHaitam El OuahabiPas encore d'évaluation
- Cours 4 MaturationDocument42 pagesCours 4 Maturationjeangredin2Pas encore d'évaluation
- Snv-Biologie Cellulaire-Les Membranes CellulairesDocument5 pagesSnv-Biologie Cellulaire-Les Membranes Cellulairesamira messoudPas encore d'évaluation
- Membrane Plasmique - WikipédiaDocument30 pagesMembrane Plasmique - Wikipédiagagnon.jonathan2007Pas encore d'évaluation
- Compartiments Intracell Et Tri Des ProtéinesDocument43 pagesCompartiments Intracell Et Tri Des Protéinesa29921Pas encore d'évaluation
- Biologie CellulaireDocument26 pagesBiologie CellulaireBourobou bourobou Chadrack gaelPas encore d'évaluation
- Cytoplasme 4Document27 pagesCytoplasme 4Oumarou KontaPas encore d'évaluation
- LysosomesDocument18 pagesLysosomesAy ManPas encore d'évaluation
- Anomalie de Transport Proteique G2Document13 pagesAnomalie de Transport Proteique G2miraPas encore d'évaluation
- Httpsstaff - Univ-Batna2.dzsitesdefaultfilesabdessemed Samirafileschapitre 11 PDFDocument12 pagesHttpsstaff - Univ-Batna2.dzsitesdefaultfilesabdessemed Samirafileschapitre 11 PDFariel.2mariemPas encore d'évaluation
- MPDFDocument4 pagesMPDFBad MaxPas encore d'évaluation
- Biosynthèse Des Lipides MembranairesDocument7 pagesBiosynthèse Des Lipides Membranaireslabidiridha8Pas encore d'évaluation
- Membrane Plasmique Partie 2Document45 pagesMembrane Plasmique Partie 2Oussama PanoramaPas encore d'évaluation
- 2.-Biosynthèse Des ProtéinesDocument65 pages2.-Biosynthèse Des Protéinesmido benPas encore d'évaluation
- Le Flux MembranaireDocument22 pagesLe Flux MembranaireYouness KhalfaouiPas encore d'évaluation
- 2 Cours Cyto Les Transports MBNDocument21 pages2 Cours Cyto Les Transports MBNegos013Pas encore d'évaluation
- Cours BCG UFR2SDocument19 pagesCours BCG UFR2SSirandou DiaoPas encore d'évaluation
- CM Biologie CellulaireDocument22 pagesCM Biologie Cellulaireleovelay34Pas encore d'évaluation
- 2 - Généralités Lipides 2Document6 pages2 - Généralités Lipides 2nour achkarPas encore d'évaluation
- 01-Enzymes Sériques Et TissulairesDocument8 pages01-Enzymes Sériques Et Tissulairesbibliotheque.masterPas encore d'évaluation
- Bcis 3Document25 pagesBcis 3rimas23rimas14Pas encore d'évaluation
- BIOTRANSFORMATIONDocument24 pagesBIOTRANSFORMATIONLok Vag100% (1)
- Polynucléaires NeutrophilesDocument9 pagesPolynucléaires Neutrophileshhaammiidd11994499Pas encore d'évaluation
- Lipides 2012Document9 pagesLipides 2012cherifPas encore d'évaluation
- U4-B01. Exploration Des Proteines Plasmatiques (Poly) - 1Document14 pagesU4-B01. Exploration Des Proteines Plasmatiques (Poly) - 1alimkettouchePas encore d'évaluation
- Cours 3. Membrane PlasmiqueDocument14 pagesCours 3. Membrane PlasmiqueNã BîlPas encore d'évaluation
- Cours Phytochimie Boumerfeg S.Document24 pagesCours Phytochimie Boumerfeg S.Clementine Eyezo'oPas encore d'évaluation
- BC .Chapitre Vi Le Syst Me EndomembranaireDocument43 pagesBC .Chapitre Vi Le Syst Me EndomembranaireNaima BouhmaidaPas encore d'évaluation
- Les Compartiments IntracellulairesDocument60 pagesLes Compartiments Intracellulairesmarie dominique patricia kouassiPas encore d'évaluation
- Généralité Sur La Membrane PlasmiqueDocument48 pagesGénéralité Sur La Membrane PlasmiqueOussama IdrissPas encore d'évaluation
- TD #5: Synthèse Des Protéines de Sécrétion Au Niveau Du REGDocument7 pagesTD #5: Synthèse Des Protéines de Sécrétion Au Niveau Du REGAy ManPas encore d'évaluation
- RESUME Biologie CellulaireDocument34 pagesRESUME Biologie CellulaireMohamed SylvestrePas encore d'évaluation
- Résumé GCDocument4 pagesRésumé GCLilia TlbPas encore d'évaluation
- COURS BC - Système EndomembranaireDocument30 pagesCOURS BC - Système EndomembranaireMohammed El MachmoulPas encore d'évaluation
- Membrane-Plasmique CompressedDocument23 pagesMembrane-Plasmique Compressedmarie dominique patricia kouassiPas encore d'évaluation
- Le Réticulum Endoplasmique - 2Document45 pagesLe Réticulum Endoplasmique - 2ilyes.jilouPas encore d'évaluation
- Chap 1 Patrtie IDocument9 pagesChap 1 Patrtie IAy IşığıPas encore d'évaluation
- 1111Cours biologie cellulaire completDocument117 pages1111Cours biologie cellulaire completrabab ouerdighiPas encore d'évaluation
- 12 PeroxysomesDocument5 pages12 PeroxysomesImen Mestar100% (1)
- TD N°2 de CytologieDocument5 pagesTD N°2 de CytologieYasmina BenPas encore d'évaluation
- Biochimie Métabolique Généralité Et AutresDocument35 pagesBiochimie Métabolique Généralité Et AutresMBAÏADOUM NGARGUINAM RodriguePas encore d'évaluation
- Cours Mod Bioch Cell Et FonctDocument73 pagesCours Mod Bioch Cell Et FonctLoubna Micha100% (1)
- 02 Revetement-CellulaireDocument6 pages02 Revetement-CellulaireSagno AgnèsPas encore d'évaluation
- Biotransformation FinaleDocument28 pagesBiotransformation FinaleIkram HamachePas encore d'évaluation
- TD DynamiqueDocument24 pagesTD Dynamiquesouttou sabrinaPas encore d'évaluation
- 02 - Revetement CellulaireDocument20 pages02 - Revetement CellulaireIssouf BerthePas encore d'évaluation
- Chapitre VII Le Reticulum Endoplasmique Et Lappareil de Golgi B8Document13 pagesChapitre VII Le Reticulum Endoplasmique Et Lappareil de Golgi B8kim namjoonPas encore d'évaluation
- Sheets Du Dossier GpoiesefDocument7 pagesSheets Du Dossier Gpoiesefyaya camaraPas encore d'évaluation
- Polycopie Voies Métaboliques S6Document42 pagesPolycopie Voies Métaboliques S6MOHSINA GHAMMATPas encore d'évaluation
- Biochimie Approfondie Cours 3Document12 pagesBiochimie Approfondie Cours 3Alhassane BahPas encore d'évaluation
- Membranes cellulaires: Les Grands Articles d'UniversalisD'EverandMembranes cellulaires: Les Grands Articles d'UniversalisPas encore d'évaluation
- Memoire Fin InfoDocument52 pagesMemoire Fin InfoMzn JannahPas encore d'évaluation
- Trafic Intracellulaire ModifiéeDocument33 pagesTrafic Intracellulaire ModifiéeMzn JannahPas encore d'évaluation
- Matrice BPC 2Document39 pagesMatrice BPC 2Mzn JannahPas encore d'évaluation
- Cours #1 - Jonction CellDocument96 pagesCours #1 - Jonction CellMzn JannahPas encore d'évaluation
- GastrulationDocument5 pagesGastrulationMinaheta Guy maurelPas encore d'évaluation
- Système MusculaireDocument7 pagesSystème MusculaireAmine WlPas encore d'évaluation
- HemostaseDocument26 pagesHemostaseLeon ChipondaPas encore d'évaluation
- TP HematologieDocument4 pagesTP HematologieSalam AbdulPas encore d'évaluation
- Devoir de Contrôle N°2 Lycée Pilote - SVT - 2ème Sciences (2011-2012) Mlle Jridi Zohra PDFDocument3 pagesDevoir de Contrôle N°2 Lycée Pilote - SVT - 2ème Sciences (2011-2012) Mlle Jridi Zohra PDFmedbhs100% (3)
- 1 - Histologie Du Tube DigestifDocument6 pages1 - Histologie Du Tube DigestifOcéane VromanPas encore d'évaluation
- Université de TananariveDocument6 pagesUniversité de TananariveAsgaraly RahimPas encore d'évaluation
- Résumé U-1 - P-2Document4 pagesRésumé U-1 - P-2ygtzdntp2j100% (1)
- Atlas de Poche - HématologieDocument203 pagesAtlas de Poche - HématologieMahefa Serge Rakotozafy100% (7)
- 2023 2024 Syllabus Option Sante LAS VdefDocument11 pages2023 2024 Syllabus Option Sante LAS VdefHelmi MzoughiPas encore d'évaluation
- Activité 4 MembraneDocument3 pagesActivité 4 Membranenews2foot offPas encore d'évaluation
- Histologie Et Cytologie de L'os NormalDocument6 pagesHistologie Et Cytologie de L'os NormalBarina100% (1)
- Pathologie Non TumoraleDocument59 pagesPathologie Non TumoraleEmmanuelPas encore d'évaluation
- 1 COURS EPITH DE REVETEMENT 2020 Polycopes-Converti PDFDocument12 pages1 COURS EPITH DE REVETEMENT 2020 Polycopes-Converti PDFrandomPas encore d'évaluation
- GRANULOPOIESEDocument48 pagesGRANULOPOIESEYingra BaraPas encore d'évaluation
- Introduction A L'anatomieDocument5 pagesIntroduction A L'anatomieYassine EssPas encore d'évaluation
- ExerciceDocument22 pagesExerciceAsgaraly Rahim0% (1)
- Histologie de La PeauDocument20 pagesHistologie de La PeauMehdiPas encore d'évaluation
- Histologie Des ReinsDocument6 pagesHistologie Des Reinsassil08Pas encore d'évaluation
- Organisation de La Cellule EucaryoteDocument2 pagesOrganisation de La Cellule EucaryoteFatma EzahraPas encore d'évaluation
- QCM de Biologie Cellulaire JMJ 2019Document4 pagesQCM de Biologie Cellulaire JMJ 2019Rym ZakraouiPas encore d'évaluation
- 129 Biologie Cellulaire Introduction Au Vivant CPCMDocument17 pages129 Biologie Cellulaire Introduction Au Vivant CPCMFleur D'anisPas encore d'évaluation
- Physio Du MuscleDocument19 pagesPhysio Du Musclesamcruz31100% (1)
- A LégenderDocument3 pagesA LégenderFatima Ezzahra AchkouPas encore d'évaluation
- Vie Cellulaire 10emeDocument7 pagesVie Cellulaire 10emeDiabyPas encore d'évaluation
- C Proc Et EucDocument23 pagesC Proc Et EucAya youyaPas encore d'évaluation
- BP 1 - La Cellule Partie 1 - CorrectionDocument3 pagesBP 1 - La Cellule Partie 1 - CorrectionBoucherie / Alimentaire DucosPas encore d'évaluation
- Developement DentDocument47 pagesDevelopement DentHasan ZaylaaPas encore d'évaluation
- Peau Et Annexes 2020Document62 pagesPeau Et Annexes 2020Ebe100% (1)
- Biologie Cellulaire. Pr. SellamiDocument35 pagesBiologie Cellulaire. Pr. SellamiNouha BoughraraPas encore d'évaluation