Vous aimerez peut-être aussi
- Se Nourrir en Conscience - Gabriel CousensDocument684 pagesSe Nourrir en Conscience - Gabriel Cousensmes réductions bioPas encore d'évaluation
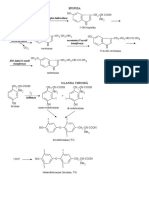
- Chimie Organique Fonctionelle S5Document120 pagesChimie Organique Fonctionelle S5أمين حسونيPas encore d'évaluation
- Fisca Internationale - Partie 2Document30 pagesFisca Internationale - Partie 2Anas Damoun100% (1)
- Aldehydes CetonesDocument29 pagesAldehydes Cetonesvaslek terriyo100% (1)
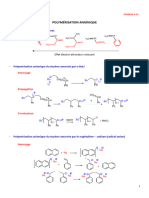
- Cours Polymères-1Document22 pagesCours Polymères-1Ouehnia tynaPas encore d'évaluation
- La Gestion Des Approvisionnements Et de StockDocument28 pagesLa Gestion Des Approvisionnements Et de Stockel khaiat mohamed aminePas encore d'évaluation
- Biologie Des Acides NucléiquesDocument22 pagesBiologie Des Acides Nucléiquesabekhti2008Pas encore d'évaluation
- Cours Chouaf - Pratiques Urbanisme Operat 2016 PDFDocument144 pagesCours Chouaf - Pratiques Urbanisme Operat 2016 PDFAhmed HoufPas encore d'évaluation
- Devis Chair PDFDocument2 pagesDevis Chair PDFHervé D'AvillaPas encore d'évaluation
- Chimie Organique Fonctionelle S5Document120 pagesChimie Organique Fonctionelle S5fskPas encore d'évaluation
- Serie TD 2 Avec Correction Glucide BiochimieDocument5 pagesSerie TD 2 Avec Correction Glucide BiochimieWissam Tiza100% (7)
- S6 HOC CH Chapitre 4Document21 pagesS6 HOC CH Chapitre 4Eddouks Fatimazahrae100% (2)
- 4° - Emprunt Et Metissage Dans La Chanson ActuelleDocument5 pages4° - Emprunt Et Metissage Dans La Chanson Actuellebamora53Pas encore d'évaluation
- Musique Chretienne Musique PaienneDocument85 pagesMusique Chretienne Musique Paiennepohrel lonkengPas encore d'évaluation
- Chapitre 2 Les Composés Phenoliques-Merghem PDFDocument22 pagesChapitre 2 Les Composés Phenoliques-Merghem PDFWahiba YahiaouiPas encore d'évaluation
- 07 MetabolismeDocument13 pages07 MetabolismeWhat are youPas encore d'évaluation
- ANTIÉPILEPTIQUESDocument35 pagesANTIÉPILEPTIQUESTidjouguena Mawena CasimirPas encore d'évaluation
- Antituberculeux PDFDocument28 pagesAntituberculeux PDFMar YemPas encore d'évaluation
- SV31 RevisionDocument10 pagesSV31 Revision李潇Pas encore d'évaluation
- Activite-Synthese D Un Edulcorant-Protection Des Fonctions OrganiquesDocument10 pagesActivite-Synthese D Un Edulcorant-Protection Des Fonctions OrganiqueshamlaouiPas encore d'évaluation
- I Modification Post TranscriptionnelleDocument25 pagesI Modification Post TranscriptionnelleFarida SirimaPas encore d'évaluation
- Chapitre 5 - PhénolsDocument8 pagesChapitre 5 - PhénolsBeatrice Florin100% (1)
- PC T ch1 BilanDocument1 pagePC T ch1 BilanZeyn NdPas encore d'évaluation
- Glucide 1Document1 pageGlucide 1imane AGOURRAMPas encore d'évaluation
- Formule HormoniDocument4 pagesFormule HormoniRalumPas encore d'évaluation
- Chimie Organique Examens Corr 05Document1 pageChimie Organique Examens Corr 05sosoukr123Pas encore d'évaluation
- TD-chimie OrgaDocument21 pagesTD-chimie OrgaJean AnanPas encore d'évaluation
- Chap7les AminesDocument4 pagesChap7les AminesChTri LyricsPas encore d'évaluation
- Classification Des HormonesDocument1 pageClassification Des HormonesIsmael100% (1)
- Chimie Orga 1Document61 pagesChimie Orga 1Badr ZemmouriPas encore d'évaluation
- Coenzymes EtDocument22 pagesCoenzymes EtLaira ImiPas encore d'évaluation
- Cours Acides Nuléique 2016Document44 pagesCours Acides Nuléique 2016Loubna EssabiiPas encore d'évaluation
- TP3-acide CinnamiqueDocument12 pagesTP3-acide CinnamiqueDenisaPas encore d'évaluation
- EX1Document6 pagesEX1zgazga amirPas encore d'évaluation
- Barbi Turi QuesDocument9 pagesBarbi Turi QuesmatPas encore d'évaluation
- Membrane PlasmiqueDocument48 pagesMembrane Plasmiqueabdalhakamrouche8Pas encore d'évaluation
- Organo PhosphoréDocument11 pagesOrgano PhosphoréZied BouabenePas encore d'évaluation
- 1.2.2-Exercices OsidesDocument2 pages1.2.2-Exercices OsidesLawrence Mundene-timothee100% (2)
- 1.acides aminés.UAHB-1Document40 pages1.acides aminés.UAHB-1paul destinPas encore d'évaluation
- Diagrammes Glucides Important PDFDocument11 pagesDiagrammes Glucides Important PDFRadhia KossaiPas encore d'évaluation
- Chapitre 6 Aldéhydes Et CétonesDocument7 pagesChapitre 6 Aldéhydes Et CétonesBeatrice Florin100% (8)
- Cours CoumarineDocument12 pagesCours CoumarineM.B. IsmailPas encore d'évaluation
- Analyse Qualitative Et Quantitative Des Anthocyanes Dans Les Produits NaturelsDocument14 pagesAnalyse Qualitative Et Quantitative Des Anthocyanes Dans Les Produits NaturelsAbdoul SawadogoPas encore d'évaluation
- Corrigé TD3Document7 pagesCorrigé TD3Mariam Ben AbdelouahabPas encore d'évaluation
- Chapitre 3 Terpenes PDFDocument10 pagesChapitre 3 Terpenes PDFmohammed safouPas encore d'évaluation
- Chapitre 1 CHM321 CoordinationDocument35 pagesChapitre 1 CHM321 CoordinationVictor NguemoPas encore d'évaluation
- Doc1 CarboxylesDocument4 pagesDoc1 CarboxylesDominique LoeuilletPas encore d'évaluation
- Signet Fonctions OrganiquesDocument1 pageSignet Fonctions OrganiquesSab 433Pas encore d'évaluation
- Respiration CellulaireDocument7 pagesRespiration Cellulaireanon-174796Pas encore d'évaluation
- TD Chimie Bio-Organique P.lerougeDocument8 pagesTD Chimie Bio-Organique P.lerougehenri.4Pas encore d'évaluation
- Master PhosphoreDocument34 pagesMaster PhosphoreMohamed EL FAGHLOUMIPas encore d'évaluation
- Acide Aminé BonDocument36 pagesAcide Aminé BonLaira ImiPas encore d'évaluation
- C9 Elemente de Biochimie PDFDocument23 pagesC9 Elemente de Biochimie PDFddeeiyaPas encore d'évaluation
- Furanne Et Dérivés-ConvertiDocument28 pagesFuranne Et Dérivés-ConvertiBel KisPas encore d'évaluation
- Intrants 7oct BertrandDocument21 pagesIntrants 7oct BertrandAL Mustapha CisséPas encore d'évaluation
- Cours Acides Nucléiques 20 21Document109 pagesCours Acides Nucléiques 20 21anassPas encore d'évaluation
- 1 Les Antiépileptiques 2017Document56 pages1 Les Antiépileptiques 2017Na FesPas encore d'évaluation
- Examen 1Document19 pagesExamen 1bchouPas encore d'évaluation
- Chapitre 4Document34 pagesChapitre 4sextansPas encore d'évaluation
- Stereochimie Chim 201-05-06cDocument13 pagesStereochimie Chim 201-05-06cToumany FofanaPas encore d'évaluation
- PC 3ème - L7 - Les AlcanesDocument9 pagesPC 3ème - L7 - Les AlcanesGedion Doua100% (1)
- Annexe Sa5Document4 pagesAnnexe Sa5Plateny AMOUSSOUPas encore d'évaluation
- Planche 4-11 - CopieDocument3 pagesPlanche 4-11 - CopieAyoub MahiPas encore d'évaluation
- td5 Chimie Bio 2006-2007Document3 pagestd5 Chimie Bio 2006-2007Anonymous dGMueaR6Pas encore d'évaluation
- Activité 4 IbuprofèneDocument4 pagesActivité 4 IbuprofèneDiego Alves SilvaPas encore d'évaluation
- Chap 4 PHYSIO LENCEPHALEDocument5 pagesChap 4 PHYSIO LENCEPHALEAnani Jacques YaoPas encore d'évaluation
- Chap 1 SYSTEME NERVEUX GENERALITESDocument7 pagesChap 1 SYSTEME NERVEUX GENERALITESAnani Jacques YaoPas encore d'évaluation
- Chapitre 1 - Généralité Sur Le Métabolisme - BIOCHIMIE METABOLIQUE ET MEDICALEDocument9 pagesChapitre 1 - Généralité Sur Le Métabolisme - BIOCHIMIE METABOLIQUE ET MEDICALEAnani Jacques YaoPas encore d'évaluation
- Objectif BiochimieDocument9 pagesObjectif BiochimieAnani Jacques YaoPas encore d'évaluation
- Chapitre 3 - Bêta-Oxydation Des Acides Gras - BIOCHIMIE METABOLIQUE POUR BM À FSSDocument9 pagesChapitre 3 - Bêta-Oxydation Des Acides Gras - BIOCHIMIE METABOLIQUE POUR BM À FSSAnani Jacques YaoPas encore d'évaluation
- DE Farines Nutrimix Togo: L'Unite Production Des AUDocument9 pagesDE Farines Nutrimix Togo: L'Unite Production Des AUAnani Jacques YaoPas encore d'évaluation
- K9C Fiches TM 39-M-15Document9 pagesK9C Fiches TM 39-M-15Matheus B. SilvaPas encore d'évaluation
- Guide Détaillé Du RZF v1.0 - 0Document30 pagesGuide Détaillé Du RZF v1.0 - 0hodjee2Pas encore d'évaluation
- Johnson, Eric - Cliffsm of DoverDocument7 pagesJohnson, Eric - Cliffsm of DoverGilvan NovaisPas encore d'évaluation
- Dans La Salle Dattente de Dieu Dans Quelle Période Êtes-VousDocument1 pageDans La Salle Dattente de Dieu Dans Quelle Période Êtes-VousdavidPas encore d'évaluation
- RapportDocument27 pagesRapportFellahi AyaPas encore d'évaluation
- Isha 5Document57 pagesIsha 5ArbnorPas encore d'évaluation
- Diagnostic Nord Ouest Nov 2019Document32 pagesDiagnostic Nord Ouest Nov 2019Roller Saint PierrePas encore d'évaluation
- Les Protocoles de Base TCPDocument17 pagesLes Protocoles de Base TCPBenie DE DieuPas encore d'évaluation
- Borderô Diário Da Anserve Data: ApartDocument18 pagesBorderô Diário Da Anserve Data: ApartKaique FreitasPas encore d'évaluation
- Ebauche Projet MemoireDocument10 pagesEbauche Projet MemoireTerry BELFLEURPas encore d'évaluation
- Support de Cours - Le Metier D'assistante de DirectionDocument17 pagesSupport de Cours - Le Metier D'assistante de DirectionDEEP 2KPas encore d'évaluation
- Pujilah Allah Dalam Bait-Nya: Panggilan IbadahDocument112 pagesPujilah Allah Dalam Bait-Nya: Panggilan IbadahJoshua ArielPas encore d'évaluation
- rabarisoaVazolyN SN M2 09Document52 pagesrabarisoaVazolyN SN M2 09RmjulienPas encore d'évaluation
- DUNGS - Pressure Switch LGW A2-A2P - Instruction ManualDocument12 pagesDUNGS - Pressure Switch LGW A2-A2P - Instruction ManualNoparit KittisatitPas encore d'évaluation
- Questions Sur La Galette Des RoisDocument2 pagesQuestions Sur La Galette Des RoisATPas encore d'évaluation
- Neuvaines Pour Le Mois D'avrilDocument1 pageNeuvaines Pour Le Mois D'avrillydiaPas encore d'évaluation
- Politique Nationale de Nutrition Et Plan National D'action Pour La Nutrition (ONN-2004)Document66 pagesPolitique Nationale de Nutrition Et Plan National D'action Pour La Nutrition (ONN-2004)HayZara Madagascar100% (1)
- MMMMM MERABET - Samira PDFDocument122 pagesMMMMM MERABET - Samira PDFMario MandžukićPas encore d'évaluation
- ReimsDocument13 pagesReimsvicnet31Pas encore d'évaluation
- Moulinex-Machine A PainDocument20 pagesMoulinex-Machine A PainLorenzzoPas encore d'évaluation
- COMPO DE PASSAGE CE2 Exp Dictee A ImprimerDocument2 pagesCOMPO DE PASSAGE CE2 Exp Dictee A Imprimerאור האהבה רוח חזקה100% (2)
- RECUEILEDE5 FinalprocidingDocument275 pagesRECUEILEDE5 FinalprocidingMansour HassanePas encore d'évaluation
- Maison Artemisia Bulletin AdhesionDocument4 pagesMaison Artemisia Bulletin AdhesionChiekhna Cheikh Sadbou FayePas encore d'évaluation