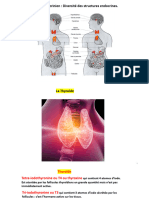
Vous aimerez peut-être aussi
- Cours Physiologie Systeme Endocri Nien 2017 2ème Année MédecineDocument161 pagesCours Physiologie Systeme Endocri Nien 2017 2ème Année Médecineanaischauvin12Pas encore d'évaluation
- Endocrinologie 1Document46 pagesEndocrinologie 1Hikari Kazue100% (1)
- 2014 7 Le Systeme EndocrinienDocument15 pages2014 7 Le Systeme EndocrinienSamira MBasskarPas encore d'évaluation
- SE Cours 3OSDocument22 pagesSE Cours 3OSLara FarquharsonPas encore d'évaluation
- EndocrinologieDocument46 pagesEndocrinologieAhmedou EllaPas encore d'évaluation
- 01 - Principes - Generaux - de - La - Physiologie - Des - Glandes - Endocrines - DR - SILINIDocument29 pages01 - Principes - Generaux - de - La - Physiologie - Des - Glandes - Endocrines - DR - SILINIkilo Romeo23Pas encore d'évaluation
- Physiologie Endocrinienne Et Métabolique.G2UEA2020Document187 pagesPhysiologie Endocrinienne Et Métabolique.G2UEA2020Mercy MwemeraPas encore d'évaluation
- Hormones GénéralitésDocument65 pagesHormones GénéralitésHadil TahraouiPas encore d'évaluation
- Cours EndocrinologieDocument192 pagesCours EndocrinologieZakariya nadirPas encore d'évaluation
- Universite Saad Dahleb Faculte de Medecine 2 Annee Medecine Cours de BiochimieDocument9 pagesUniversite Saad Dahleb Faculte de Medecine 2 Annee Medecine Cours de BiochimieDîãmønd QùeenPas encore d'évaluation
- Physiologie Endocrine Partie 1Document36 pagesPhysiologie Endocrine Partie 1cep100% (1)
- 1-Generalites en EndocrinoDocument21 pages1-Generalites en EndocrinobasassynenedianePas encore d'évaluation
- Cours Endocrinologie de La ReproductionDocument49 pagesCours Endocrinologie de La ReproductionLukusa MauricePas encore d'évaluation
- Cours N°1d'endocrinologie S4 2020Document32 pagesCours N°1d'endocrinologie S4 2020Younes Masoud100% (1)
- 1-Generalites en Endocrinologie PDFDocument21 pages1-Generalites en Endocrinologie PDFthierno amadou bahPas encore d'évaluation
- P Fonctions Du Système Endocrinien - (L. Bayer) - 23.04.21Document8 pagesP Fonctions Du Système Endocrinien - (L. Bayer) - 23.04.21nasten16fhPas encore d'évaluation
- Généralités Sur La Physiologie Du Système EndocrinienDocument5 pagesGénéralités Sur La Physiologie Du Système EndocrinienSaken667Pas encore d'évaluation
- Cours 6Document27 pagesCours 6BouabidiPas encore d'évaluation
- Chapitre 3Document67 pagesChapitre 3Widad El barhoumiPas encore d'évaluation
- Endocrinologie 23 1 2022Document53 pagesEndocrinologie 23 1 2022DA4OUR ytPas encore d'évaluation
- Physiologie Endocrinienne C01Document45 pagesPhysiologie Endocrinienne C01amael saint-louisPas encore d'évaluation
- Poly S1 PCEM2 2020Document113 pagesPoly S1 PCEM2 2020pvfbszf9mtPas encore d'évaluation
- Cours Dendocrinologie 2021-2022 - Partie 1Document32 pagesCours Dendocrinologie 2021-2022 - Partie 1carineyenkeyPas encore d'évaluation
- Pharmacologie Endocrinienne 01Document185 pagesPharmacologie Endocrinienne 01Ždn ÂśšiaPas encore d'évaluation
- 01-Principes Des Systemes de Controle HormonalDocument2 pages01-Principes Des Systemes de Controle HormonalLem AhPas encore d'évaluation
- UE8 TC Physio Cours 2 - Régulation Endocrinienne Tutorat 2013-2014Document10 pagesUE8 TC Physio Cours 2 - Régulation Endocrinienne Tutorat 2013-2014Ali EL Kihel100% (1)
- Le Système EndocrinienDocument23 pagesLe Système Endocrinienchadi ouechtatiPas encore d'évaluation
- Endocrinologie IDocument88 pagesEndocrinologie Iritajbeily1611Pas encore d'évaluation
- Cours - 3 Les Régulation Hormonale de La MétabolismeDocument9 pagesCours - 3 Les Régulation Hormonale de La Métabolismemiss anthropocenePas encore d'évaluation
- Principes Des Systemes de Controle HormonalDocument2 pagesPrincipes Des Systemes de Controle Hormonalfafoufa133Pas encore d'évaluation
- Chap Iv Systeme EndocrinienDocument56 pagesChap Iv Systeme Endocrinienbaazizitopo mohamedPas encore d'évaluation
- 1.système EndocrinienDocument31 pages1.système EndocrinienSid ahmed Zerrouki sbaPas encore d'évaluation
- CM 7 PhysioDocument31 pagesCM 7 PhysioMehdiPas encore d'évaluation
- 1 Principes Du Système de Contrôle Hormonal BDocument61 pages1 Principes Du Système de Contrôle Hormonal BOXGAMINGXOPas encore d'évaluation
- Endocrinologie 2024Document95 pagesEndocrinologie 2024mohamedazizhmida000Pas encore d'évaluation
- Version ImprimableDocument22 pagesVersion ImprimableSylvain GravelaisPas encore d'évaluation
- Physio Endocrino23-01glandes EndocrinesDocument4 pagesPhysio Endocrino23-01glandes EndocrinesVictorin FANDREBOPas encore d'évaluation
- Résumé GCDocument4 pagesRésumé GCLilia TlbPas encore d'évaluation
- Fiche TD Signalisation Cellulaire L3 BPA S6Document7 pagesFiche TD Signalisation Cellulaire L3 BPA S6Massouh Assoui100% (3)
- Cours Physiologie S4 2017Document62 pagesCours Physiologie S4 2017Narciso RodriguezPas encore d'évaluation
- Cours 3Document16 pagesCours 3azizhseck2Pas encore d'évaluation
- QE de Physiologie EndocrinoDocument12 pagesQE de Physiologie EndocrinoN.Pas encore d'évaluation
- EndocrinotoxicitéDocument82 pagesEndocrinotoxicitéKhaoula Kouloughli100% (1)
- Cours Histologie Glandes Endocrines 1Document3 pagesCours Histologie Glandes Endocrines 1Dany MelPas encore d'évaluation
- 1 Endocrinologie GeneraleDocument74 pages1 Endocrinologie GeneraleHq sishPas encore d'évaluation
- TD1 Regulation SABRINA.2019Document5 pagesTD1 Regulation SABRINA.2019axelPas encore d'évaluation
- L'axe Hypothalamo-HypophysaireDocument11 pagesL'axe Hypothalamo-Hypophysairegeorge.guergourPas encore d'évaluation
- 01-HORMONES GénéralitésDocument14 pages01-HORMONES GénéralitésalimkettouchePas encore d'évaluation
- Endocrinologie s6 Dahbi.-1Document9 pagesEndocrinologie s6 Dahbi.-1Zineb DahmaniPas encore d'évaluation
- Ahh M2decineDocument22 pagesAhh M2decinemalakzerizer2005Pas encore d'évaluation
- LE SYSTÈME ENDOCRINIEN Anatomie PhysiologieDocument38 pagesLE SYSTÈME ENDOCRINIEN Anatomie PhysiologieIkramo CrPas encore d'évaluation
- Physiologie Animale Cours 8Document52 pagesPhysiologie Animale Cours 8g69wwpfw5kPas encore d'évaluation
- Le Systeme Endocrinien WDocument9 pagesLe Systeme Endocrinien WsafemindPas encore d'évaluation
- Principes Des Systèmes de Contrôle HormonalDocument34 pagesPrincipes Des Systèmes de Contrôle HormonalSouha MessaoudiPas encore d'évaluation
- 1-Physiologie Endocrinienne Generalites en EndocrinologieDocument102 pages1-Physiologie Endocrinienne Generalites en EndocrinologieAfdal BiosicaPas encore d'évaluation
- Cours EM Fusionné 2020Document99 pagesCours EM Fusionné 2020Serge Mbainoudji TendoPas encore d'évaluation
- EndocrinologieDocument43 pagesEndocrinologiePierre MonharoulPas encore d'évaluation
- Révisions 2.2Document16 pagesRévisions 2.2lenapaquiePas encore d'évaluation
- 5.0 Endocrinologie NaassilaDocument62 pages5.0 Endocrinologie NaassilaMohamed AraouchiPas encore d'évaluation
- Hormones: Les Grands Articles d'UniversalisD'EverandHormones: Les Grands Articles d'UniversalisPas encore d'évaluation
- Protéines MedecineDocument10 pagesProtéines MedecineZiko FathiPas encore d'évaluation
- Nitin Trivedi Et AlDocument7 pagesNitin Trivedi Et AlMohamed CHADLIPas encore d'évaluation
- Techniques D'immobilisation Des Cellules Et Leur ApplicationDocument50 pagesTechniques D'immobilisation Des Cellules Et Leur Applicationhouhou_houhou83% (24)
- EnzymologieDocument5 pagesEnzymologieMor GanitePas encore d'évaluation
- ThA Se HALDocument149 pagesThA Se HALAnonymous MKSfyYyODPPas encore d'évaluation
- Propriétés Générales Des EnzymesDocument28 pagesPropriétés Générales Des EnzymesliliPas encore d'évaluation
- Cazorla G. ... Planification Programmation Et Périodisation de L'entraînement 1Document44 pagesCazorla G. ... Planification Programmation Et Périodisation de L'entraînement 1Rait AitPas encore d'évaluation
- Biochimie2 PDFDocument2 pagesBiochimie2 PDFTCHEMMIK NazihaPas encore d'évaluation
- SUBSTRATS CARBONES INDUSTRIELS 8 PagesDocument8 pagesSUBSTRATS CARBONES INDUSTRIELS 8 Pagesfellah aminaPas encore d'évaluation
- Cours Cinétique Enzymatique. 21docxDocument7 pagesCours Cinétique Enzymatique. 21docxKEBIR ZAHRAPas encore d'évaluation
- Foie Medicaments2020sahliDocument6 pagesFoie Medicaments2020sahli123Pas encore d'évaluation
- Régulation Du Métabolisme Glucidique - ÉtudiantsDocument48 pagesRégulation Du Métabolisme Glucidique - ÉtudiantsfafoulolPas encore d'évaluation
- Fermentations Industrielles 10-11Document45 pagesFermentations Industrielles 10-11Hasnaa Mb100% (1)
- Orniformation Probatoire 2007 D SVTDocument8 pagesOrniformation Probatoire 2007 D SVTmoivoici147Pas encore d'évaluation
- DS N°3 25-11-23 Métabolisme 2nde - CorrectionDocument4 pagesDS N°3 25-11-23 Métabolisme 2nde - Correctionninostampa1Pas encore d'évaluation
- Mpungi K Genie BiochimieDocument84 pagesMpungi K Genie BiochimieBeevens KijiPas encore d'évaluation
- Paces Ac Ue1 BBM 15 21Document151 pagesPaces Ac Ue1 BBM 15 21Jul AzardiPas encore d'évaluation
- Cours MRE SORODocument21 pagesCours MRE SOROAlexandre OuffouePas encore d'évaluation
- Casimir Blonski - EnzymologieDocument35 pagesCasimir Blonski - Enzymologiealphonse1988Pas encore d'évaluation
- Cinétique EnzymatiqueDocument14 pagesCinétique Enzymatiqueرزقي إبن فلسطينPas encore d'évaluation
- Les Biocapteurs Enzymatiques: République Algérienne Démocratique Et PopulaireDocument15 pagesLes Biocapteurs Enzymatiques: République Algérienne Démocratique Et PopulaireLezghed Amira meriamPas encore d'évaluation
- Cours de Biologie Les EnzymesDocument10 pagesCours de Biologie Les EnzymesOussamaPas encore d'évaluation
- Examen de Cin Et Cat Avec Correction Session Normale 2018-2019Document6 pagesExamen de Cin Et Cat Avec Correction Session Normale 2018-2019Chai Mae100% (1)
- Cours Chimie Verte 2012Document110 pagesCours Chimie Verte 2012partiraretirapasPas encore d'évaluation
- 3202210312-U3Document11 pages3202210312-U3MLAN HesnaPas encore d'évaluation
- Entraînement Physique Et Sportif: Mise Au PointDocument25 pagesEntraînement Physique Et Sportif: Mise Au PointMoha CinPas encore d'évaluation
- Pal & PacDocument14 pagesPal & PacDoha HrouchPas encore d'évaluation
- Cours Bior Acteurs CHEBLI 2020Document65 pagesCours Bior Acteurs CHEBLI 2020Aghai MohamedPas encore d'évaluation
- Chapitre 3Document22 pagesChapitre 3MariemBoujmalPas encore d'évaluation
- Ferrary 2019Document10 pagesFerrary 2019Sam KhemPas encore d'évaluation