Vous aimerez peut-être aussi
- 1 Introduction A La Physiologie Des Glandes EndocrinesDocument16 pages1 Introduction A La Physiologie Des Glandes EndocrinesAlina Topor Psi-MedicalPas encore d'évaluation
- Anatomie Humaine: Un Manuel Pratique et Intuitif pour Découvrir le Corps Humain et Toutes ses ComposantesD'EverandAnatomie Humaine: Un Manuel Pratique et Intuitif pour Découvrir le Corps Humain et Toutes ses ComposantesPas encore d'évaluation
- Physiologie Endocrine Partie 1Document36 pagesPhysiologie Endocrine Partie 1cep100% (1)
- Cours IFCS 1° Année 2011Document39 pagesCours IFCS 1° Année 2011Fatim-Zahra DrissiPas encore d'évaluation
- UE8 TC Physio Cours 2 - Régulation Endocrinienne Tutorat 2013-2014Document10 pagesUE8 TC Physio Cours 2 - Régulation Endocrinienne Tutorat 2013-2014Ali EL Kihel100% (1)
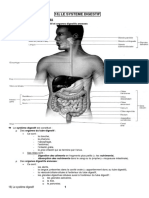
- Fonction DigestiveDocument22 pagesFonction DigestiveHamza SerPas encore d'évaluation
- 3 APPAREIL GENITAL FEMININ WordDocument13 pages3 APPAREIL GENITAL FEMININ WordFaïsole Adimou Ola100% (2)
- Le Systeme Endocrinien Cours Universite 2011Document35 pagesLe Systeme Endocrinien Cours Universite 2011Billy Bel Vocale MesidorPas encore d'évaluation
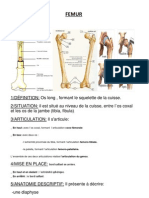
- FemurDocument4 pagesFemurBesma SbPas encore d'évaluation
- Groupes Aliments PDFDocument5 pagesGroupes Aliments PDFOtmane ELPas encore d'évaluation
- Cycle MenstruelDocument15 pagesCycle MenstruelBarinaPas encore d'évaluation
- L'appareil ReproducteurDocument118 pagesL'appareil ReproducteurAlexiaFoderaPas encore d'évaluation
- B5-Système DigestifDocument41 pagesB5-Système DigestifCléanePas encore d'évaluation
- Exploration de La CorticosurrénaleDocument58 pagesExploration de La CorticosurrénaleHicham BenaoumPas encore d'évaluation
- Glande SurrénaleDocument4 pagesGlande SurrénalehichemPas encore d'évaluation
- 07 Les NeurotransmetteursDocument20 pages07 Les NeurotransmetteursBook HunterPas encore d'évaluation
- O Esophagi TeDocument33 pagesO Esophagi TeRamdane RamdanoPas encore d'évaluation
- Hyper HypothyroïdiesDocument97 pagesHyper HypothyroïdiesBaz BazPas encore d'évaluation
- Le Systeme Endocrinien WDocument9 pagesLe Systeme Endocrinien WsafemindPas encore d'évaluation
- Cours de Biochimie MétaboliqueDocument108 pagesCours de Biochimie MétaboliqueMBAÏADOUM NGARGUINAM Rodrigue100% (1)
- Médicaments Du Système DigestifDocument137 pagesMédicaments Du Système DigestifAlexandre Kpangny Béni100% (1)
- 1ere S2 Biochimie - Vitamine Et Coenzyme-2Document70 pages1ere S2 Biochimie - Vitamine Et Coenzyme-2Majd Mahfoudh100% (1)
- AnemieDocument1 pageAnemieSam Oum Mehdi100% (1)
- Physiologie de L'équilibre Acide-BaseDocument31 pagesPhysiologie de L'équilibre Acide-BaseSOLTANIPas encore d'évaluation
- Pancréatite AigueDocument24 pagesPancréatite Aiguebina NDOYEPas encore d'évaluation
- Biophysique II GroujDocument42 pagesBiophysique II GroujAdonis Serghini100% (1)
- Résumé QE Physio-1Document20 pagesRésumé QE Physio-1Elidlbi IbrahimPas encore d'évaluation
- 1.troubles de Cycles Et de ReglesDocument48 pages1.troubles de Cycles Et de Reglespapinou sarrPas encore d'évaluation
- TD N5. Exempl Corr.Document8 pagesTD N5. Exempl Corr.Mehdi Si-HassenPas encore d'évaluation
- Les NeurotransmétteursDocument5 pagesLes NeurotransmétteursFaculté De Médecine Béchar100% (1)
- Livre Blanc de La Nutrition KalenjiDocument31 pagesLivre Blanc de La Nutrition KalenjiThę Kïñg100% (2)
- Pathologie Cellulaire Et Tissulaire - Lésions Élémentaires 2019FMP PDFDocument91 pagesPathologie Cellulaire Et Tissulaire - Lésions Élémentaires 2019FMP PDFYounes AbidPas encore d'évaluation
- Hydrocéphalie DR SamaiDocument6 pagesHydrocéphalie DR SamaiYou SraPas encore d'évaluation
- GoutteDocument20 pagesGoutteTahraoui Omar100% (1)
- 6eme Séance - 2Document42 pages6eme Séance - 2Mohamed AlouanePas encore d'évaluation
- Secretion PancreatiqueDocument15 pagesSecretion PancreatiqueZahia ArichaPas encore d'évaluation
- Physiologie Du ColonDocument33 pagesPhysiologie Du Colonyaya camaraPas encore d'évaluation
- Cours 6Document27 pagesCours 6BouabidiPas encore d'évaluation
- Les Compartiments LiquidiensDocument14 pagesLes Compartiments LiquidiensFati ZoraPas encore d'évaluation
- Cours D3 GoutteDocument11 pagesCours D3 GouttePierre SchlaederPas encore d'évaluation
- Milieu Intérieur 2Document23 pagesMilieu Intérieur 2Ons AouinetPas encore d'évaluation
- Physiologie Des Cycles MenstruelsDocument10 pagesPhysiologie Des Cycles MenstruelsNabil holmes100% (1)
- Automatisme CardiaqueDocument58 pagesAutomatisme CardiaqueyvesPas encore d'évaluation
- Appareil Génital MaleDocument49 pagesAppareil Génital MaleTreycia PamboPas encore d'évaluation
- TD Calcul Des Doses Et DébitsDocument25 pagesTD Calcul Des Doses Et DébitsEya TalbiPas encore d'évaluation
- 241 Hypothyroïdie - 0Document2 pages241 Hypothyroïdie - 0Duy LêPas encore d'évaluation
- Le Système ImmunitaireDocument18 pagesLe Système ImmunitaireccPas encore d'évaluation
- Anatomie VasculaireDocument10 pagesAnatomie VasculairemerymorenaaPas encore d'évaluation
- Infections UrinairesDocument20 pagesInfections UrinairesAllo DocteurPas encore d'évaluation
- 03 LE GLOBULE ROUGE Cours 4eme Année Pharmacie DR ABBASSENDocument4 pages03 LE GLOBULE ROUGE Cours 4eme Année Pharmacie DR ABBASSENAziz BenachourPas encore d'évaluation
- 1 Acide BaseDocument4 pages1 Acide BaseSalifou Abdoul Nasser100% (1)
- 5 Les Annexes EmbryonnairesDocument12 pages5 Les Annexes EmbryonnairesBook Hunter100% (2)
- Pharm09 Bioch-Exploration Acides AminesDocument48 pagesPharm09 Bioch-Exploration Acides AminesHicham BenaoumPas encore d'évaluation
- Massage Cardiaque ExterneDocument7 pagesMassage Cardiaque ExterneZZakiiPas encore d'évaluation
- Physiologie de La Thyroide 2018Document46 pagesPhysiologie de La Thyroide 2018تقوى اللهPas encore d'évaluation
- Physio DigestiveDocument90 pagesPhysio DigestiveKnizou Afif100% (1)
- Syndrome MétaboliqueDocument10 pagesSyndrome MétaboliqueYounes TechlabPas encore d'évaluation
- Biochimie de L'inflammationDocument49 pagesBiochimie de L'inflammationsafemind100% (1)
- Les Cours Des Infirmiers en Toutes Les Modules - ANATOMIE ET PHYSIOLOGIE DE L'APPAREIL DIGESTIFDocument7 pagesLes Cours Des Infirmiers en Toutes Les Modules - ANATOMIE ET PHYSIOLOGIE DE L'APPAREIL DIGESTIFNathan DurandPas encore d'évaluation
- Acides Nucleiques PDFDocument90 pagesAcides Nucleiques PDFZineb DahmaniPas encore d'évaluation
- Sommaire: in Vitro Des Acides Nucléiques .............................................. 111Document2 pagesSommaire: in Vitro Des Acides Nucléiques .............................................. 111Zineb DahmaniPas encore d'évaluation
- 1-Structure Bases, Nucléotides Et Acides Nucléiques ARN/ADNDocument8 pages1-Structure Bases, Nucléotides Et Acides Nucléiques ARN/ADNZineb DahmaniPas encore d'évaluation
- Diversite Mesures PDFDocument7 pagesDiversite Mesures PDFZineb DahmaniPas encore d'évaluation
- EIE - ASS - Chem-Def-V3.1 - Web 19-Mars-2014 PDFDocument179 pagesEIE - ASS - Chem-Def-V3.1 - Web 19-Mars-2014 PDFZineb DahmaniPas encore d'évaluation
- MicrobiologieDocument17 pagesMicrobiologieZineb DahmaniPas encore d'évaluation
- MicrobiologieDocument64 pagesMicrobiologieZineb DahmaniPas encore d'évaluation
- Support TD Génétique - S5 - 2017 - 2018 - PR Achtak PDFDocument8 pagesSupport TD Génétique - S5 - 2017 - 2018 - PR Achtak PDFZineb Dahmani50% (2)
- Croissance Bacterienne PDFDocument6 pagesCroissance Bacterienne PDFZineb DahmaniPas encore d'évaluation
- Tspé-ENERGIE-chapitre 3 Contraction Musculaire Et ATP Prof PDFDocument3 pagesTspé-ENERGIE-chapitre 3 Contraction Musculaire Et ATP Prof PDFZineb Dahmani100% (1)
- Diaporama Respiration Chapitre 4-5 PDFDocument30 pagesDiaporama Respiration Chapitre 4-5 PDFZineb DahmaniPas encore d'évaluation
- TD7 Les Organites de La Cellule PDFDocument42 pagesTD7 Les Organites de La Cellule PDFZineb DahmaniPas encore d'évaluation
- n-1 PDFDocument39 pagesn-1 PDFZineb DahmaniPas encore d'évaluation
- 6 0018 17 PDFDocument71 pages6 0018 17 PDFZineb DahmaniPas encore d'évaluation
- EIE - ASS - Chem-Def-V3.1 - Web 19-Mars-2014 PDFDocument179 pagesEIE - ASS - Chem-Def-V3.1 - Web 19-Mars-2014 PDFZineb DahmaniPas encore d'évaluation
- Articlesebtegoula2016 PDFDocument15 pagesArticlesebtegoula2016 PDFZineb DahmaniPas encore d'évaluation
- Phonetique de Francais PDFDocument122 pagesPhonetique de Francais PDFZineb Dahmani100% (3)
- Document PDFDocument206 pagesDocument PDFZineb DahmaniPas encore d'évaluation
- Cours sv4 Flor Msanda1 p16Document27 pagesCours sv4 Flor Msanda1 p16Zineb DahmaniPas encore d'évaluation
- Cours Biochimie Lipides SV3Document65 pagesCours Biochimie Lipides SV3Zineb DahmaniPas encore d'évaluation
- Ranunculaceae Brassicaceae MalvaceaeDocument37 pagesRanunculaceae Brassicaceae MalvaceaeZineb DahmaniPas encore d'évaluation
- Génétique 1 - Partie 1Document43 pagesGénétique 1 - Partie 1Zineb DahmaniPas encore d'évaluation
- Série D - ExercicesDocument5 pagesSérie D - ExercicesZineb Dahmani40% (5)
- Cours Prot+®ines 2016Document43 pagesCours Prot+®ines 2016Zineb DahmaniPas encore d'évaluation
- Prévenir Les Tms Chez SoignantsDocument15 pagesPrévenir Les Tms Chez SoignantsMerieme SafaaPas encore d'évaluation
- Information IRMDocument1 pageInformation IRMVictor DolcePas encore d'évaluation
- Les Substances Bioactives Dans Les Fruits Et LagumesDocument6 pagesLes Substances Bioactives Dans Les Fruits Et LagumesJosue Lubilanji BamuinamunaPas encore d'évaluation
- Bilan Des Consultations Specialisees en Medecine Du TravailDocument76 pagesBilan Des Consultations Specialisees en Medecine Du TravailBienfait KabwikaPas encore d'évaluation
- Devoir de Contrôle N°1 - SVT Reproduction - Bac Sciences Exp (2010-2011) MR Obey JobraneDocument3 pagesDevoir de Contrôle N°1 - SVT Reproduction - Bac Sciences Exp (2010-2011) MR Obey JobraneamineabidiPas encore d'évaluation
- Placenta PraeviaDocument34 pagesPlacenta PraeviaINESPas encore d'évaluation
- Carcinome Epidermoide Cutane Rapport IntegralDocument110 pagesCarcinome Epidermoide Cutane Rapport IntegralBek GhizlanePas encore d'évaluation
- La Fin Des MaladiesDocument93 pagesLa Fin Des MaladiesdelemerPas encore d'évaluation
- La PasteurisationDocument16 pagesLa PasteurisationGedion Doua100% (1)
- Brochure M1 PPCP 2020 2021Document55 pagesBrochure M1 PPCP 2020 2021Gabriela AviñaPas encore d'évaluation
- 05-Presentation Deflechies POLYCOPEDocument19 pages05-Presentation Deflechies POLYCOPEOussama MamouniPas encore d'évaluation
- Alexandre Jodorowsky - La Voie de L'imaginationDocument480 pagesAlexandre Jodorowsky - La Voie de L'imaginationabdelbassiryoungaPas encore d'évaluation
- Brouillon Exposé de Communication IIDocument1 pageBrouillon Exposé de Communication IISarrPas encore d'évaluation
- Mieuxvivre2023 s1 GrossesseDocument184 pagesMieuxvivre2023 s1 GrossessecisnerosPas encore d'évaluation
- Planchage Radiologie Ostéo-Articulaire 12 (Rachis) AC CTDocument88 pagesPlanchage Radiologie Ostéo-Articulaire 12 (Rachis) AC CTRafik LaghPas encore d'évaluation
- Le Manuel Du Résident Neurologie 2017 PDFDocument3 467 pagesLe Manuel Du Résident Neurologie 2017 PDFTo Ma Du BwaPas encore d'évaluation
- Conduite À Tenir Devant Une Dermatose Bulleuse de LDocument7 pagesConduite À Tenir Devant Une Dermatose Bulleuse de Lserignemodou100% (1)
- Concepts de Base en Psychologie Analytique 1Document8 pagesConcepts de Base en Psychologie Analytique 1Cyber OsloPas encore d'évaluation
- H28. Niger - Rapport - Formation Des Enquêteurs Sur Le Travail Forcé - NewDocument9 pagesH28. Niger - Rapport - Formation Des Enquêteurs Sur Le Travail Forcé - Newcign bnetdPas encore d'évaluation
- Rapport de Stage ElsaDocument17 pagesRapport de Stage ElsaRosé Ibrahim OkePas encore d'évaluation
- Guidepsh1fr 064679500 1525 26062017Document16 pagesGuidepsh1fr 064679500 1525 26062017assoumoudenis42Pas encore d'évaluation
- Pas 96 2014 FoodDocument44 pagesPas 96 2014 FoodAbdeljalil OUIDANIPas encore d'évaluation
- Les Tumeurs Intracanalaires Papillaires Et Mucineuses Du Pancreas Multifocales: Prise en Charge Et Alternatives Therapeutiques PossiblesDocument9 pagesLes Tumeurs Intracanalaires Papillaires Et Mucineuses Du Pancreas Multifocales: Prise en Charge Et Alternatives Therapeutiques PossiblesIJAR JOURNALPas encore d'évaluation
- 01 - Pied-GeneralitesDocument91 pages01 - Pied-Generalitesguemiri lyesPas encore d'évaluation
- Evaluation Sec 06Document2 pagesEvaluation Sec 06Barthélemy HoubenPas encore d'évaluation
- FT Amplix Neisseria Gonorrhoeae - FR Ang - V1Document13 pagesFT Amplix Neisseria Gonorrhoeae - FR Ang - V1Fouad RahiouyPas encore d'évaluation
- Les Conséquences de La Porno DépendanceDocument5 pagesLes Conséquences de La Porno DépendanceJean-LucPas encore d'évaluation
- Odonto 2021 RemionDocument87 pagesOdonto 2021 Remionlina rayenePas encore d'évaluation
- Dictée Préparée 5èmeDocument21 pagesDictée Préparée 5èmeBakaye DembelePas encore d'évaluation
- Fluidothérapie Cerba+Vet+CollegeDocument20 pagesFluidothérapie Cerba+Vet+CollegechatonblibluPas encore d'évaluation