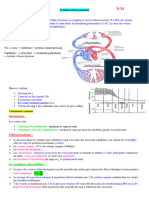
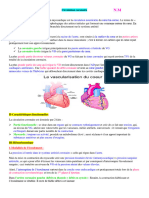
Vous aimerez peut-être aussi
- GENETIQUE FONDAMENTALE - Transcription - MoodleDocument3 pagesGENETIQUE FONDAMENTALE - Transcription - MoodleGh7Pas encore d'évaluation
- 11 Traduction Chap IV Expression Inf GenDocument5 pages11 Traduction Chap IV Expression Inf GenRock In BandPas encore d'évaluation
- Cours BMDocument22 pagesCours BMNidhaloo NidPas encore d'évaluation
- Cours BMGG 2018Document43 pagesCours BMGG 2018Fares ManelPas encore d'évaluation
- 16-Traduction de L'information GénétiqueDocument33 pages16-Traduction de L'information GénétiqueInsafPas encore d'évaluation
- Les Ribosomes Et La Synthèse Protéique B6Document10 pagesLes Ribosomes Et La Synthèse Protéique B6kim namjoonPas encore d'évaluation
- Med Genetique-Transcription EucaryotesDocument20 pagesMed Genetique-Transcription EucaryotesRacha MalakPas encore d'évaluation
- Biologie Moléculaire Chapitre 3 REGULATION M1 1Document8 pagesBiologie Moléculaire Chapitre 3 REGULATION M1 1اميرة في عرش ابيPas encore d'évaluation
- 3 Synthèse Protéique-2Document12 pages3 Synthèse Protéique-2Sagacious IvejutenPas encore d'évaluation
- Chapitre 12 Synthèse Des Protéines (Transcription-Traduction)Document5 pagesChapitre 12 Synthèse Des Protéines (Transcription-Traduction)One Love LifePas encore d'évaluation
- HAV504B - Chapitre 2Document24 pagesHAV504B - Chapitre 2Ylona RamondPas encore d'évaluation
- Transcription 21 - 11 - 2023Document55 pagesTranscription 21 - 11 - 2023Elmahdi BOUHFIDPas encore d'évaluation
- 6-Transcription Et Maturation Des ARNDocument10 pages6-Transcription Et Maturation Des ARNmed.hmamouch99Pas encore d'évaluation
- Bio Mol Cours 12Document10 pagesBio Mol Cours 12Abdelghaffar EL-AMMARIPas encore d'évaluation
- 3eme Biochimie 2019-2020 Mahdab N L3 Bioch COURS DE BIO MOLDocument22 pages3eme Biochimie 2019-2020 Mahdab N L3 Bioch COURS DE BIO MOLry yukPas encore d'évaluation
- Ziani Module Biologie Moléculaire Et Génie Génétique L3 MicobiologieDocument122 pagesZiani Module Biologie Moléculaire Et Génie Génétique L3 MicobiologieOwassiPas encore d'évaluation
- Biologie Moléculaire Chapitre 5 M1 1Document5 pagesBiologie Moléculaire Chapitre 5 M1 1اميرة في عرش ابيPas encore d'évaluation
- UE2 - Biologie Cellulaire - Le CytosolDocument11 pagesUE2 - Biologie Cellulaire - Le Cytosolsun-nee-chan9Pas encore d'évaluation
- La Synthèse Des ProtéinesDocument3 pagesLa Synthèse Des Protéinesbouchakour meryem100% (4)
- Protéines RecombinantesDocument10 pagesProtéines RecombinantesYahia Berbouchi100% (1)
- 1 TranscriptionDocument12 pages1 TranscriptionYounes YouyouPas encore d'évaluation
- BG3 ExosDocument22 pagesBG3 Exospersonne12Pas encore d'évaluation
- Biologie Moléculaire Chapitre 1 T Procaryote M1Document8 pagesBiologie Moléculaire Chapitre 1 T Procaryote M1اميرة في عرش ابيPas encore d'évaluation
- Chapitre Syntha Se ProtaiqueDocument29 pagesChapitre Syntha Se ProtaiqueNouha HadjPas encore d'évaluation
- Cours RéglationDocument33 pagesCours RéglationBADzPas encore d'évaluation
- Synthèses Des ProtéinesDocument1 pageSynthèses Des ProtéinesAl sarahPas encore d'évaluation
- Evaluation 2Document5 pagesEvaluation 2firdaoussaoud22Pas encore d'évaluation
- Le REDocument6 pagesLe RENimabPas encore d'évaluation
- I. Définition: Les Ribosomes Ou Grains de Palade SontDocument3 pagesI. Définition: Les Ribosomes Ou Grains de Palade Sontnouralc.2005Pas encore d'évaluation
- Cours 8 - La Synthèse ProtéiqueDocument8 pagesCours 8 - La Synthèse ProtéiqueFarouk Ben YahiaPas encore d'évaluation
- Activité 7 Traduction Chapitre 1 Unité 2Document3 pagesActivité 7 Traduction Chapitre 1 Unité 2hamamielloPas encore d'évaluation
- Objectif Terminal: Expliquer Les Différents Mécanismes Intervenants Dans La Synthèse Des ProtéinesDocument2 pagesObjectif Terminal: Expliquer Les Différents Mécanismes Intervenants Dans La Synthèse Des ProtéinesBenji JeanPas encore d'évaluation
- Système Endomembranaire - NotesDocument18 pagesSystème Endomembranaire - Notesemiliedurosoir2Pas encore d'évaluation
- SVT 1ère C - L6 - La Synthèse Des ProtéinesDocument16 pagesSVT 1ère C - L6 - La Synthèse Des ProtéinesNamoryPas encore d'évaluation
- Bio Mol L3 MicrobiologieDocument18 pagesBio Mol L3 MicrobiologieimanePas encore d'évaluation
- TraductionDocument36 pagesTraductionnonamethe427Pas encore d'évaluation
- Le RibosomeDocument38 pagesLe Ribosomeabdalhakamrouche8Pas encore d'évaluation
- Biologie Moléculaire Et GG 2Document30 pagesBiologie Moléculaire Et GG 2Sangaré soungaloPas encore d'évaluation
- A Synthèse Des ProtéinesDocument3 pagesA Synthèse Des ProtéinesNadaPas encore d'évaluation
- Traduction L2 SADocument33 pagesTraduction L2 SAiliazPas encore d'évaluation
- TRANSCRIPTION S4 ÉtudiantsDocument6 pagesTRANSCRIPTION S4 ÉtudiantsNiavo HeriniriantsoaPas encore d'évaluation
- Chapitre 03Document5 pagesChapitre 03You NesPas encore d'évaluation
- BCM 7 Transcription TraductionDocument56 pagesBCM 7 Transcription TraductionJoel AmaniPas encore d'évaluation
- CHEIDIDocument4 pagesCHEIDIcheilla nandjioPas encore d'évaluation
- La Transcription l3 Microbiologie 2022Document8 pagesLa Transcription l3 Microbiologie 2022rou röuPas encore d'évaluation
- Ed Biomol 2Document2 pagesEd Biomol 2Alexia HritcuPas encore d'évaluation
- Duplication de l'ADN - 011009Document65 pagesDuplication de l'ADN - 011009nonamethe427Pas encore d'évaluation
- TRAnscriptionDocument16 pagesTRAnscriptionmaysounPas encore d'évaluation
- Série d'exercices N°1 - SVT expression de l'I G, genigénétique et mutation - 3ème Sciences exp (2023-2024) Mr مصباح العربيDocument4 pagesSérie d'exercices N°1 - SVT expression de l'I G, genigénétique et mutation - 3ème Sciences exp (2023-2024) Mr مصباح العربيkadra55amri100% (1)
- BiochDocument243 pagesBiochyouness.khalfaouiPas encore d'évaluation
- Genetique1an Transcription Adn2017hanachiDocument37 pagesGenetique1an Transcription Adn2017hanachisoumeya soltaniPas encore d'évaluation
- Exposé Du Groupe 9Document35 pagesExposé Du Groupe 9Grâce KABOREPas encore d'évaluation
- 6.NOUVEAU LE Réticulum Endoplasmique Et Les Ribosomes 2022Document68 pages6.NOUVEAU LE Réticulum Endoplasmique Et Les Ribosomes 2022Le BoyaïstePas encore d'évaluation
- Epissage Chez Les EucaryotesDocument6 pagesEpissage Chez Les EucaryotesAlain Bertrand JuniorPas encore d'évaluation
- CM04 - Traduction Et Routage Des ProteinesDocument8 pagesCM04 - Traduction Et Routage Des ProteinesRock In BandPas encore d'évaluation
- Cours 2 MTDocument65 pagesCours 2 MTحسن غالية حسن غاليةPas encore d'évaluation
- Genetique1an16 TranscriptionDocument10 pagesGenetique1an16 TranscriptionRacha MalakPas encore d'évaluation
- .Expression Du Génome Humain - PCEM2 - IB - Septembre 2022Document100 pages.Expression Du Génome Humain - PCEM2 - IB - Septembre 2022oumaimahenteti5Pas encore d'évaluation
- SVT TA - L7 - La Biosynthèse Des ProtéinesDocument12 pagesSVT TA - L7 - La Biosynthèse Des Protéinestangaramariam566Pas encore d'évaluation
- La TranscriptionDocument28 pagesLa TranscriptionMelissa BaloulPas encore d'évaluation
- Syndromes TopographiquesDocument30 pagesSyndromes TopographiquesSafaa MedPas encore d'évaluation
- Syndromes BulbairesDocument8 pagesSyndromes Bulbairesbrahim brahimPas encore d'évaluation
- MPR Lombalgie ImagerieDocument11 pagesMPR Lombalgie ImagerieHOUSSEYN GHOZLANEPas encore d'évaluation
- Infections UrinairesDocument25 pagesInfections UrinairesHOUSSEYN GHOZLANEPas encore d'évaluation
- Epuration Extra RénaleDocument48 pagesEpuration Extra RénaleHOUSSEYN GHOZLANEPas encore d'évaluation
- ŒdèmesDocument35 pagesŒdèmesHOUSSEYN GHOZLANEPas encore d'évaluation
- 4-Transport Des Gaz Dans Le Sang N.MDocument6 pages4-Transport Des Gaz Dans Le Sang N.MHOUSSEYN GHOZLANEPas encore d'évaluation
- 5-Système À Basse Pression N.MDocument3 pages5-Système À Basse Pression N.MHOUSSEYN GHOZLANEPas encore d'évaluation
- 5 - Régulation RespiratoireDocument14 pages5 - Régulation RespiratoireHOUSSEYN GHOZLANEPas encore d'évaluation
- 1-Introduction À La Physiologie Respiratoire N.MDocument3 pages1-Introduction À La Physiologie Respiratoire N.MHOUSSEYN GHOZLANEPas encore d'évaluation
- 6-Circulation Coronaire N.MDocument3 pages6-Circulation Coronaire N.MHOUSSEYN GHOZLANEPas encore d'évaluation
- Confusion Mentale Cours WordDocument10 pagesConfusion Mentale Cours WordHOUSSEYN GHOZLANEPas encore d'évaluation
- Troubles Anxieux Et TOCDocument18 pagesTroubles Anxieux Et TOCHOUSSEYN GHOZLANEPas encore d'évaluation
- 09.09 Hematomes CerebrauxDocument49 pages09.09 Hematomes CerebrauxHOUSSEYN GHOZLANEPas encore d'évaluation
- Les Addictions PrésentationDocument24 pagesLes Addictions PrésentationHOUSSEYN GHOZLANEPas encore d'évaluation
- 11 - IRC de L - Adulte PR ARZOURDocument46 pages11 - IRC de L - Adulte PR ARZOURHOUSSEYN GHOZLANEPas encore d'évaluation
- Urgences PsychiatriquesDocument10 pagesUrgences PsychiatriquesHOUSSEYN GHOZLANEPas encore d'évaluation
- 10irp02 Fiche Memo Obesite Adulte Bilan Initial D Un Exces de PoidsDocument1 page10irp02 Fiche Memo Obesite Adulte Bilan Initial D Un Exces de PoidsHOUSSEYN GHOZLANEPas encore d'évaluation
- Troubles BipolairesDocument20 pagesTroubles BipolairesHOUSSEYN GHOZLANEPas encore d'évaluation
- Extraction de LDocument13 pagesExtraction de LMirabilis MinoPas encore d'évaluation
- Biochimie Approfondie Cours 7Document25 pagesBiochimie Approfondie Cours 7Alhassane BahPas encore d'évaluation
- College Protestant Comp. Tle D - 074611Document2 pagesCollege Protestant Comp. Tle D - 074611GeremyPas encore d'évaluation
- CoursDocument16 pagesCoursKoki kokiPas encore d'évaluation
- TD1 2019Document3 pagesTD1 2019Cheikh ThiamPas encore d'évaluation
- TP 011Document2 pagesTP 011maysounPas encore d'évaluation
- Travaux Diriges de Genetique PHM 110 2022Document18 pagesTravaux Diriges de Genetique PHM 110 2022Remadji vieriPas encore d'évaluation
- Interactions Cellulaires Dans Le FoieDocument8 pagesInteractions Cellulaires Dans Le Foiemaxence tricaudPas encore d'évaluation
- La Thérapie Génique À La Rescousse de La Maladie D'alzheimerDocument2 pagesLa Thérapie Génique À La Rescousse de La Maladie D'alzheimerMila Vargas PlacenciaPas encore d'évaluation
- Les Mécanismes de La Méiose - MaxicoursDocument8 pagesLes Mécanismes de La Méiose - MaxicoursJean HovozounkouPas encore d'évaluation
- Série de Génétique Des DiploïdesDocument6 pagesSérie de Génétique Des DiploïdesAdem MechriPas encore d'évaluation
- Modes de Transmission Hereditaire Des Caracteres MonofactorielsDocument9 pagesModes de Transmission Hereditaire Des Caracteres MonofactorielsMariettePas encore d'évaluation
- Polycopie Du Cours Régulation Des Gènes Chez Les Procaryotes M1 Génétique MM GHARZOULIDocument64 pagesPolycopie Du Cours Régulation Des Gènes Chez Les Procaryotes M1 Génétique MM GHARZOULISarah SisiPas encore d'évaluation
- Corps Humain 2H UE 2.2 2017Document43 pagesCorps Humain 2H UE 2.2 2017Nambinintsoa HeriniainaPas encore d'évaluation
- Hipraviar+trt4 Af TN 713459 00.0Document1 pageHipraviar+trt4 Af TN 713459 00.0Mohamed BachaPas encore d'évaluation
- Evaluation Des Activites de Vaccination Dans Le District Sanitaire de MacinaDocument91 pagesEvaluation Des Activites de Vaccination Dans Le District Sanitaire de MacinaA PPas encore d'évaluation
- Caryotype TK Et IndicationsDocument65 pagesCaryotype TK Et IndicationsahmedzPas encore d'évaluation
- Topo VHB 2021Document46 pagesTopo VHB 2021gourariPas encore d'évaluation
- Staphylococcus Aureus: Résistante À La PénicillineDocument2 pagesStaphylococcus Aureus: Résistante À La PénicillineMeena Machado-MucchielliPas encore d'évaluation
- Techniques ImmunologiquesDocument1 pageTechniques ImmunologiquesqalbonPas encore d'évaluation
- Exercices Biologie Moleculaire PDFDocument2 pagesExercices Biologie Moleculaire PDFMike50% (10)
- References-Histoire PharmacieDocument3 pagesReferences-Histoire PharmacieSaulo CarneiroPas encore d'évaluation
- Dépliant Séminaire SNBPMS 2024-2-1Document4 pagesDépliant Séminaire SNBPMS 2024-2-1Moh Nadjib RebiziPas encore d'évaluation
- Cahier D'exercice3Document12 pagesCahier D'exercice3GHERMI .M80% (15)
- TP N°5 (1ère S)Document2 pagesTP N°5 (1ère S)sandra.le.du100% (2)
- Revision Reproduction Humaine CorrigeDocument29 pagesRevision Reproduction Humaine CorrigeDavid KingPas encore d'évaluation
- TD BHS 263 PDFDocument2 pagesTD BHS 263 PDFPaul Mbede Sedena100% (1)
- Le Développement Des Biotechnologies: Quelques Repères HistoriquesDocument14 pagesLe Développement Des Biotechnologies: Quelques Repères HistoriquesmostecocPas encore d'évaluation
- COURS GENETIQUE (CHAPITRE 1,2,3,4,5,6,7, Et 8) 2019-2020Document70 pagesCOURS GENETIQUE (CHAPITRE 1,2,3,4,5,6,7, Et 8) 2019-2020laurentkalambayi850Pas encore d'évaluation
- 1er Cours Introduction Au Monde MicrobienDocument8 pages1er Cours Introduction Au Monde MicrobienMoufida Trénitarya100% (1)