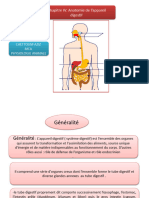
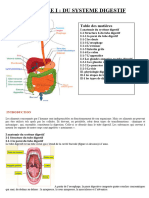
Vous aimerez peut-être aussi
- Hormones: Les Grands Articles d'UniversalisD'EverandHormones: Les Grands Articles d'UniversalisPas encore d'évaluation
- MetaboDocument78 pagesMetaboBaryyon XPas encore d'évaluation
- Muscles: Les Grands Articles d'UniversalisD'EverandMuscles: Les Grands Articles d'UniversalisPas encore d'évaluation
- OesophageDocument4 pagesOesophageLolo KawtchiPas encore d'évaluation
- Physiologie DigestiveDocument40 pagesPhysiologie DigestiveGregory GebertPas encore d'évaluation
- Physiologie de la digestion _BIOS_3066Document40 pagesPhysiologie de la digestion _BIOS_3066steve fotsoPas encore d'évaluation
- L'Appareil Digestif - 1Document19 pagesL'Appareil Digestif - 1El FaroukPas encore d'évaluation
- Anatomie Et Physiologie Du Système DigestifDocument8 pagesAnatomie Et Physiologie Du Système Digestifaurelien choupoPas encore d'évaluation
- Résumé Digestion Biologie, Spécialité: NutritionDocument18 pagesRésumé Digestion Biologie, Spécialité: NutritionFatima Lydia GacemPas encore d'évaluation
- TD N5. Exempl Corr.Document8 pagesTD N5. Exempl Corr.Mehdi Si-HassenPas encore d'évaluation
- Histologie de L'appareil DigestifDocument15 pagesHistologie de L'appareil DigestiftessPas encore d'évaluation
- L'appareil Digestif: 1-Structure Générale Du Tube DigestifDocument8 pagesL'appareil Digestif: 1-Structure Générale Du Tube DigestifSunset ShimmerPas encore d'évaluation
- Cours Physiologie DigestiveDocument21 pagesCours Physiologie DigestiveAlaa RmidaPas encore d'évaluation
- Anatomie Physiologie Digestive1Document13 pagesAnatomie Physiologie Digestive1Jo NoutPas encore d'évaluation
- 21 - LestomacDocument5 pages21 - Lestomacsameh SakaPas encore d'évaluation
- Cours de Digestion (S5)Document12 pagesCours de Digestion (S5)Houssam ChebliPas encore d'évaluation
- L'estomac: Dochead Sous-DocheadDocument9 pagesL'estomac: Dochead Sous-DocheadJJPas encore d'évaluation
- Unité 6 L'appareil DegistifDocument66 pagesUnité 6 L'appareil DegistifMery ChristmasPas encore d'évaluation
- chapitre IV physiologie de système digestifDocument44 pageschapitre IV physiologie de système digestifRomario NkomoPas encore d'évaluation
- Team-J HISTO SPECIALDocument13 pagesTeam-J HISTO SPECIALroger kinaPas encore d'évaluation
- Physiologie de La Mastication Et SalivationDocument7 pagesPhysiologie de La Mastication Et SalivationChaima benPas encore d'évaluation
- Histo de L'app DigestifDocument20 pagesHisto de L'app DigestifÏk RamPas encore d'évaluation
- 05 - Notes - DigestifDocument54 pages05 - Notes - DigestifAgathe MassantPas encore d'évaluation
- Physiologie DigestiveDocument48 pagesPhysiologie DigestiveYouness Khalfaoui75% (4)
- SPLANCHNOLOGIEDocument6 pagesSPLANCHNOLOGIEdhcbhvct7bPas encore d'évaluation
- B5-Système DigestifDocument41 pagesB5-Système DigestifCléanePas encore d'évaluation
- 1.généralité Sur La Physiologie DegistiveDocument13 pages1.généralité Sur La Physiologie DegistiveBer MouPas encore d'évaluation
- Physiologie Du Système Gastro-Intestinal 2018-2019-COURS ENTIER-Pr WATCHODocument44 pagesPhysiologie Du Système Gastro-Intestinal 2018-2019-COURS ENTIER-Pr WATCHOlucienntakobajira1Pas encore d'évaluation
- 1-Généralités Sur La Physiologie DigestiveDocument7 pages1-Généralités Sur La Physiologie DigestiveBouch BousmPas encore d'évaluation
- Resumé de La Physiologie Digestive, Par Oussama Keddad PDFDocument41 pagesResumé de La Physiologie Digestive, Par Oussama Keddad PDFsifouPas encore d'évaluation
- CHAP 2 - Sys DigestifDocument20 pagesCHAP 2 - Sys DigestifThibault LongreePas encore d'évaluation
- Physiologie DigestiveDocument27 pagesPhysiologie Digestiveyaya camaraPas encore d'évaluation
- 5-La Paroi GastriqueDocument10 pages5-La Paroi GastriqueSifo GuendouzPas encore d'évaluation
- Physiologie Digestive 4Document75 pagesPhysiologie Digestive 4Marcus MilitoPas encore d'évaluation
- Cours Syst Digertifs Et Respiratoire Pour IUA DOUALA 2023-2024Document19 pagesCours Syst Digertifs Et Respiratoire Pour IUA DOUALA 2023-2024jessicangambongPas encore d'évaluation
- Ľ Appariel Dijestif + Ľ Appariel GenitalDocument28 pagesĽ Appariel Dijestif + Ľ Appariel GenitalAyoub LilouPas encore d'évaluation
- 5-Le ColonDocument6 pages5-Le Colonkilo Romeo23Pas encore d'évaluation
- EyamoDocument5 pagesEyamoEyamo.LydiePas encore d'évaluation
- Appareil DigestifDocument36 pagesAppareil DigestifYoucef HoualefPas encore d'évaluation
- Le Gros Intestin V 2016-2017Document2 pagesLe Gros Intestin V 2016-2017Pace MakerPas encore d'évaluation
- Polycopié Cours S5 Digestion PHy GRan FOn 2021 PDFDocument13 pagesPolycopié Cours S5 Digestion PHy GRan FOn 2021 PDFSalma SalmaaaPas encore d'évaluation
- Physiologie - Digestive ''''Document52 pagesPhysiologie - Digestive ''''Manassé MisengePas encore d'évaluation
- Appareil Digestif 2ème Année Véto Cours 1 2021Document73 pagesAppareil Digestif 2ème Année Véto Cours 1 2021walid bb100% (1)
- Colon Et Canal AnalDocument4 pagesColon Et Canal AnalMohamed FerniPas encore d'évaluation
- Traité en PhysiologieDocument8 pagesTraité en PhysiologieAlhassane BahPas encore d'évaluation
- Appareil DigestifDocument16 pagesAppareil DigestifAdn AdnPas encore d'évaluation
- Power AnapatheDocument16 pagesPower Anapatheilham ZaimechePas encore d'évaluation
- Cours Tissus St2sDocument9 pagesCours Tissus St2sDieuboue ArthurPas encore d'évaluation
- Les Annélides #Document1 pageLes Annélides #enora filleurPas encore d'évaluation
- Physio Animal Système_digestifDocument127 pagesPhysio Animal Système_digestifomarimohamad06Pas encore d'évaluation
- UE 2.2 Endoc, Digestif Et Uro TutoratDocument27 pagesUE 2.2 Endoc, Digestif Et Uro TutoratlouaPas encore d'évaluation
- Physiologie Tractus DigestifDocument16 pagesPhysiologie Tractus DigestifCharlottePas encore d'évaluation
- S2chap4coursTDPhyioDigest1 45Document45 pagesS2chap4coursTDPhyioDigest1 45Noura AzaouagPas encore d'évaluation
- BCH 306 - Tube Digestif Et FoieDocument27 pagesBCH 306 - Tube Digestif Et FoieEmma NtiPas encore d'évaluation
- Physiologie DigestiveDocument49 pagesPhysiologie DigestiveAnou KalengaPas encore d'évaluation
- EXPOSEDocument44 pagesEXPOSERania Prya MUTHUSAMY RENGASAMY100% (1)
- 1-La Phase Bucco-Œsophagienne de La Digestion.1362058756Document56 pages1-La Phase Bucco-Œsophagienne de La Digestion.1362058756DëLux OwOPas encore d'évaluation
- Histologie de Lintestin GreleDocument6 pagesHistologie de Lintestin Grelegodwe ernestPas encore d'évaluation
- Appareil Digestif Humain 1Document6 pagesAppareil Digestif Humain 1Hritcu CatalinaPas encore d'évaluation
- Physio Digest L1S2 Ziguinch 2022Document13 pagesPhysio Digest L1S2 Ziguinch 2022antaafall464Pas encore d'évaluation
- Maghat Cours Chimie Generale SVT-S1 2021-2022Document84 pagesMaghat Cours Chimie Generale SVT-S1 2021-2022Youssef RguiguePas encore d'évaluation
- TD Genetique 2014Document6 pagesTD Genetique 2014Youssef RguiguePas encore d'évaluation
- Roles Des Éléments MinérauxDocument10 pagesRoles Des Éléments MinérauxYoussef RguiguePas encore d'évaluation
- RésperationDocument46 pagesRésperationYoussef RguiguePas encore d'évaluation
- Olivier 2005Document9 pagesOlivier 2005mehdiPas encore d'évaluation
- Magnet MED Notice INSECTICIDEDocument2 pagesMagnet MED Notice INSECTICIDEYoussef RguiguePas encore d'évaluation
- Memoire Julien LANOE DUASDocument119 pagesMemoire Julien LANOE DUASVanPas encore d'évaluation
- C'Est de L'eau ChansonDocument1 pageC'Est de L'eau ChansonThomas NicolePas encore d'évaluation
- 117328-Article Text-325356-1-10-20150520Document6 pages117328-Article Text-325356-1-10-20150520Mohamed Abdellaoui AbdellaouiPas encore d'évaluation
- MAROUA 3eDocument20 pagesMAROUA 3eFranck KenduPas encore d'évaluation
- 2 Documents Sciences Les Volcans2Document12 pages2 Documents Sciences Les Volcans2KLEINPas encore d'évaluation
- Forçage Radiatif Et Conséquences Enseignement Scientifique TerminaleDocument3 pagesForçage Radiatif Et Conséquences Enseignement Scientifique TerminalehttpssoatePas encore d'évaluation
- Le Grand Livre Des Monsieur Madame Hargreaves 1934 Annas ArchiveDocument172 pagesLe Grand Livre Des Monsieur Madame Hargreaves 1934 Annas ArchiveMenziwokuhle DladlaPas encore d'évaluation
- TABLEAU RECAPITULATIF DES FREQUENCES DES DESORDRES EN FONCTION DES FACIERS - Copie (3) - CopieDocument3 pagesTABLEAU RECAPITULATIF DES FREQUENCES DES DESORDRES EN FONCTION DES FACIERS - Copie (3) - CopieRIKOPas encore d'évaluation
- 1 Notions-FondamentalesDocument136 pages1 Notions-FondamentalesIlyass ZerqtouniPas encore d'évaluation
- Metodo Arban para Trombon 01Document15 pagesMetodo Arban para Trombon 01Pablo Gustavo Letona100% (1)
- Rapport TechniqueDocument1 pageRapport TechniqueAbdelkader BOULADJEMAR100% (1)
- VulclimDocument23 pagesVulclimvalentyPas encore d'évaluation
- PDF Francais EssentielDocument8 pagesPDF Francais EssentielAmine AlichePas encore d'évaluation
- Les VannesDocument2 pagesLes Vannesbyrnndi worldPas encore d'évaluation
- Controle À Domicile - Semestre 1 - 2APIC - 2021-2022Document2 pagesControle À Domicile - Semestre 1 - 2APIC - 2021-2022Hello HelloPas encore d'évaluation
- 2CTC435058D1701Document1 page2CTC435058D1701RafiurRahmanPas encore d'évaluation
- Flore Vegetation MediterraneennesDocument74 pagesFlore Vegetation Mediterraneenneshamid_touhamiPas encore d'évaluation
- Protections CoaxialesDocument1 pageProtections CoaxialesEl Saiid TojoorPas encore d'évaluation
- Devoir1 2Document4 pagesDevoir1 2badrPas encore d'évaluation
- ConductionDocument6 pagesConductionThanina BelkouchePas encore d'évaluation
- Ecoulement Turbulent-Cas D'une GrilleDocument20 pagesEcoulement Turbulent-Cas D'une GrillenouhailaPas encore d'évaluation
- E 040105Document7 pagesE 040105Kassim MdiPas encore d'évaluation
- ExamenBia2002 PDFDocument22 pagesExamenBia2002 PDFaymenPas encore d'évaluation
- Bon Resume ECG PDFDocument3 pagesBon Resume ECG PDFChanez FekirPas encore d'évaluation
- Chap 4 - Mã©thodes SismiquesDocument45 pagesChap 4 - Mã©thodes Sismiquesntimeh BoulanPas encore d'évaluation
- Cours Hydrologie GénéraleDocument306 pagesCours Hydrologie GénéraleAhmed Ahmeed64% (11)
- 2 Le Climat Global de La Terre Et Son ÉvolutionDocument5 pages2 Le Climat Global de La Terre Et Son ÉvolutionGevantika TharmaseelanPas encore d'évaluation
- Eolienne 1Document70 pagesEolienne 1amine bounounaPas encore d'évaluation
- Declaration Du VendeurDocument5 pagesDeclaration Du Vendeurhugo737977Pas encore d'évaluation
- Geologie Revisions 5emeDocument4 pagesGeologie Revisions 5emeAkram ́Pas encore d'évaluation
- Microbiologie médicale I: agents pathogènes et microbiome humainD'EverandMicrobiologie médicale I: agents pathogènes et microbiome humainÉvaluation : 5 sur 5 étoiles5/5 (1)
- Quelle alimentation pour moi ?: Régime sans cholestérol, sans gluten, femme enceinte...D'EverandQuelle alimentation pour moi ?: Régime sans cholestérol, sans gluten, femme enceinte...Pas encore d'évaluation
- Atlas de neuroanatomie fonctionnelle: Première édition françaiseD'EverandAtlas de neuroanatomie fonctionnelle: Première édition françaisePas encore d'évaluation
- Régime Cétogène: Le Guide Ultime Pour Maîtriser Une Perte De Poids Saine (Recettes De Régime Céto)D'EverandRégime Cétogène: Le Guide Ultime Pour Maîtriser Une Perte De Poids Saine (Recettes De Régime Céto)Pas encore d'évaluation
- Equilibre Hormonal: Comment regagner l’équilibre d’hormones, la pulsion sexuelle, le sommeil et perdre du poids dès maintenantD'EverandEquilibre Hormonal: Comment regagner l’équilibre d’hormones, la pulsion sexuelle, le sommeil et perdre du poids dès maintenantPas encore d'évaluation