Vous aimerez peut-être aussi
- Symboliques Du Corps HumainDocument20 pagesSymboliques Du Corps HumainWat Brans100% (3)
- L Électricité À La MaisonDocument15 pagesL Électricité À La MaisonLrachidPas encore d'évaluation
- 26.05.pneumopathies D'hypersensibilitéDocument16 pages26.05.pneumopathies D'hypersensibilitéIna GrecuPas encore d'évaluation
- Pneumo4an-Abces Poumon MessadiDocument10 pagesPneumo4an-Abces Poumon MessadiMi RyPas encore d'évaluation
- Rapport de Stage de Pédiatrie DéfinitifDocument53 pagesRapport de Stage de Pédiatrie DéfinitifTarik83% (6)
- Les Heritiers - 3 - La Prison Doree - Erin WattDocument378 pagesLes Heritiers - 3 - La Prison Doree - Erin WattNaomie Marciano100% (2)
- Allergie et hypersensibilité: Les Grands Articles d'UniversalisD'EverandAllergie et hypersensibilité: Les Grands Articles d'UniversalisPas encore d'évaluation
- Fiche de PosteDocument2 pagesFiche de PosteMed ChemliPas encore d'évaluation
- Pneumologie (Golden)Document264 pagesPneumologie (Golden)Islam ShakhabovPas encore d'évaluation
- Electrolyse Cours 2008 PDFDocument5 pagesElectrolyse Cours 2008 PDFAliMchirguiPas encore d'évaluation
- Tous allergiques ?: Comprendre une épidémie contemporaineD'EverandTous allergiques ?: Comprendre une épidémie contemporainePas encore d'évaluation
- L'asthme de L' - eDocument8 pagesL'asthme de L' - eWini PuatiPas encore d'évaluation
- Rhinite Chronique Non AllergiqueDocument20 pagesRhinite Chronique Non Allergiqueammar eidPas encore d'évaluation
- 94 - AsthmeDocument94 pages94 - AsthmeSofia RhellabPas encore d'évaluation
- Plantier Lasthme PharmacieDocument45 pagesPlantier Lasthme PharmaciehuiPas encore d'évaluation
- AsthmeDocument12 pagesAsthmeMeriem BoulenouarPas encore d'évaluation
- Fiche LiSA-Hypersensibilité Et Allergies Respiratoires Chez L'enfant Et Chez L'aDocument11 pagesFiche LiSA-Hypersensibilité Et Allergies Respiratoires Chez L'enfant Et Chez L'aaugustinemorelPas encore d'évaluation
- 5-L'ASTHME - CopieDocument22 pages5-L'ASTHME - CopieOueslati KhalilPas encore d'évaluation
- PNP EosinophilesDocument30 pagesPNP EosinophilesLoulou popPas encore d'évaluation
- Asthme Enfant Nourrisson 1ere PartieDocument9 pagesAsthme Enfant Nourrisson 1ere PartieFlaminusPas encore d'évaluation
- 10 AsthmeDocument11 pages10 AsthmeKhaled SghaierPas encore d'évaluation
- Asthme 1Document35 pagesAsthme 1Amira NourPas encore d'évaluation
- Physiopathologie de L'asthmeDocument28 pagesPhysiopathologie de L'asthmeKheira RahouiPas encore d'évaluation
- Tes HipertensiDocument3 pagesTes HipertensiNormilaPas encore d'évaluation
- Le Cas Clinique Du Mois - Aspergillose Pulm - Un Cas d'ASP Nécrosante Chronique - Golinval - 07 - RevMedLgDocument3 pagesLe Cas Clinique Du Mois - Aspergillose Pulm - Un Cas d'ASP Nécrosante Chronique - Golinval - 07 - RevMedLgandybalebuamagambuPas encore d'évaluation
- Rhinite AllergiqueDocument6 pagesRhinite AllergiquehalilounanafaizaPas encore d'évaluation
- Asthme de L'enfant Et Du Nourrisson-ConvertiDocument13 pagesAsthme de L'enfant Et Du Nourrisson-ConvertiFeriel AePas encore d'évaluation
- Cours Pneumo - Med4 - 2018Document107 pagesCours Pneumo - Med4 - 2018Mahamadou WAGUEPas encore d'évaluation
- Expose 4 Plantes Medecinales-3Document84 pagesExpose 4 Plantes Medecinales-3hossein kouahiPas encore d'évaluation
- La Toux 2010Document7 pagesLa Toux 2010Éric Comlan SewaiPas encore d'évaluation
- L'asthme Fiche Resumé CompleteDocument5 pagesL'asthme Fiche Resumé Completeclaralucchese06Pas encore d'évaluation
- 8.5.rhinite Non AllergiqueDocument9 pages8.5.rhinite Non AllergiquenonoPas encore d'évaluation
- La PollinoseDocument2 pagesLa PollinoseB.nacerPas encore d'évaluation
- Les TDocument131 pagesLes TdjaballahPas encore d'évaluation
- LeconimprimDocument6 pagesLeconimprimAnouamane AlantiPas encore d'évaluation
- Asthme de L'adulteDocument26 pagesAsthme de L'adulteJasmy FfPas encore d'évaluation
- Module 4 La Pneumonie Aigue Communaitaire PDFDocument49 pagesModule 4 La Pneumonie Aigue Communaitaire PDFA BPas encore d'évaluation
- 18 - Asthme - AdultDocument6 pages18 - Asthme - AdultGhassan ShehabPas encore d'évaluation
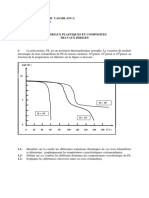
- 4.6.1 Mesure de L - Inflammation Bronchique (Resultats)Document11 pages4.6.1 Mesure de L - Inflammation Bronchique (Resultats)yousratouileb99Pas encore d'évaluation
- Lupus 2023Document63 pagesLupus 2023fatimamourghi50Pas encore d'évaluation
- Les Pneumopathies Interstitielles Diffuses Idiopathiques RochatDocument6 pagesLes Pneumopathies Interstitielles Diffuses Idiopathiques RochatMichael LjuslinPas encore d'évaluation
- Physiopathologie de L'AsthmeDocument13 pagesPhysiopathologie de L'AsthmeHenry TraoréPas encore d'évaluation
- Méningite Aseptique Due Aux AINS Chez LupiqueDocument3 pagesMéningite Aseptique Due Aux AINS Chez LupiqueHenry TraoréPas encore d'évaluation
- BronsectaziiDocument6 pagesBronsectaziiCristina LupascoPas encore d'évaluation
- La TouxDocument33 pagesLa TouxGasmi MostafaPas encore d'évaluation
- Toux Chronique de Ladulte CEP Cours 2010Document20 pagesToux Chronique de Ladulte CEP Cours 2010Michel RamanantsoavinaPas encore d'évaluation
- L AsthmeDocument14 pagesL AsthmeNour El Houda MKPas encore d'évaluation
- Asthme Du Nourrisson Et de L'enfantDocument44 pagesAsthme Du Nourrisson Et de L'enfantvague2000Pas encore d'évaluation
- Partie Du Diagnostic Et Traitement: COVID-19 Prévention Et Le Contrôle DesDocument12 pagesPartie Du Diagnostic Et Traitement: COVID-19 Prévention Et Le Contrôle DesKiassa Med HaithemPas encore d'évaluation
- L'asthmeDocument26 pagesL'asthmeOuattara BamoryPas encore d'évaluation
- 8 AsthmeDocument4 pages8 AsthmeSoli TairPas encore d'évaluation
- Fiche LiSA-Infections Broncho Pulmonaires Communautaires de L'adulte Et de L'enfant - LiSADocument37 pagesFiche LiSA-Infections Broncho Pulmonaires Communautaires de L'adulte Et de L'enfant - LiSAjustelauraPas encore d'évaluation
- EosinophilieDocument12 pagesEosinophilieikou ikramPas encore d'évaluation
- Les BronchitesDocument15 pagesLes BronchitesNouha ChellaliPas encore d'évaluation
- Asthme Et TraitementDocument36 pagesAsthme Et TraitementHoussem EddinePas encore d'évaluation
- Choc Anaphylactique - ClinicalKeyDocument27 pagesChoc Anaphylactique - ClinicalKeyBilal AfkirPas encore d'évaluation
- Infections Récurrentes Des Voies Respiratoires Inférieures Chez L'enfantDocument80 pagesInfections Récurrentes Des Voies Respiratoires Inférieures Chez L'enfantBa abkPas encore d'évaluation
- Asthme Professionnelle Cours Pour Etudiants 6eme Annee DR Bensekhria PDFDocument5 pagesAsthme Professionnelle Cours Pour Etudiants 6eme Annee DR Bensekhria PDFzakisami2008Pas encore d'évaluation
- Asthme Bronchique PR F ChaoukiDocument56 pagesAsthme Bronchique PR F ChaoukiAbdourasck GOUMANEH100% (1)
- Fiche LiSA-Infections Broncho Pulmonaires Communautaires de L'adulte Et de L'enfDocument17 pagesFiche LiSA-Infections Broncho Pulmonaires Communautaires de L'adulte Et de L'enfaugustinemorelPas encore d'évaluation
- Main 2Document8 pagesMain 2Koké Lamine TraoréPas encore d'évaluation
- Fichier Produit 3019Document18 pagesFichier Produit 3019Mamadou TandiaPas encore d'évaluation
- Asthme Du Sujet Âgé FinalDocument68 pagesAsthme Du Sujet Âgé FinalahmadranauiPas encore d'évaluation
- Diagnostic Des Encéphalites Autoimmunes: Version of RecordDocument28 pagesDiagnostic Des Encéphalites Autoimmunes: Version of RecordWafae TouchaPas encore d'évaluation
- Troubles de L'oculomotricité: RésuméDocument12 pagesTroubles de L'oculomotricité: RésuméNabil BouhafaraPas encore d'évaluation
- Tachycardies Ventriculaires: B Brembilla-PerrotDocument13 pagesTachycardies Ventriculaires: B Brembilla-PerrotNabil BouhafaraPas encore d'évaluation
- Indications de L'arthroscopie Du Poignet: RésuméDocument4 pagesIndications de L'arthroscopie Du Poignet: RésuméNabil BouhafaraPas encore d'évaluation
- Méthodes Non Traumatiques D'évaluation de La Densité Minérale Et de La Structure OsseuseDocument8 pagesMéthodes Non Traumatiques D'évaluation de La Densité Minérale Et de La Structure OsseuseNabil BouhafaraPas encore d'évaluation
- Notice Zeclar 250mg Comp. Pelli. B 10Document2 pagesNotice Zeclar 250mg Comp. Pelli. B 10Nabil BouhafaraPas encore d'évaluation
- Serie AlcoolDocument5 pagesSerie AlcoolPFE100% (2)
- Notice Jeulin Cellule de MalassezDocument7 pagesNotice Jeulin Cellule de MalassezMalo AubryPas encore d'évaluation
- Gestion & Adm Du Pers.Document76 pagesGestion & Adm Du Pers.Innocent BIAPas encore d'évaluation
- CyberPower NI DS BU600-1000E FR FR v2Document2 pagesCyberPower NI DS BU600-1000E FR FR v2Kada Ben youcefPas encore d'évaluation
- BUPHA T 2013 HEYMONET CLAUDE Inflamation GeneralitéDocument200 pagesBUPHA T 2013 HEYMONET CLAUDE Inflamation GeneralitéIkram SamidPas encore d'évaluation
- Hager Techn-Katalog H3+ FRDocument180 pagesHager Techn-Katalog H3+ FRGregoire BoutouyouPas encore d'évaluation
- La Réforme Des Accidents de Travail VFDocument33 pagesLa Réforme Des Accidents de Travail VFhamza benbrekPas encore d'évaluation
- FR B2 Cpto PR J 2019Document11 pagesFR B2 Cpto PR J 2019MarianoPas encore d'évaluation
- Toitures & CharpentesDocument21 pagesToitures & CharpentesGueye AlimatouPas encore d'évaluation
- FR en de Es Ru NL It PL: Gysmi 80P - 130P 160P - 200PDocument56 pagesFR en de Es Ru NL It PL: Gysmi 80P - 130P 160P - 200PMichel GiustinaPas encore d'évaluation
- 1 Bac Les Reactions Acido Basiques Cours 1Document6 pages1 Bac Les Reactions Acido Basiques Cours 1rihabPas encore d'évaluation
- Dermatologie, Guide PratiqueDocument66 pagesDermatologie, Guide PratiqueAmélie GarceauPas encore d'évaluation
- Syllabus NégociationDocument5 pagesSyllabus NégociationechcharradiassiaPas encore d'évaluation
- 1211 PDFDocument28 pages1211 PDFHenderson TshikwataPas encore d'évaluation
- Savel Naomi 58651800 2019-2020Document66 pagesSavel Naomi 58651800 2019-2020message poor vuPas encore d'évaluation
- TP Onduleur MonophaseDocument14 pagesTP Onduleur MonophaseSouhaib LoudaPas encore d'évaluation
- Polio ReferenceDocument24 pagesPolio ReferenceRochelle CionPas encore d'évaluation
- Ais Gestion Des Ressources Hydriques BbaDocument12 pagesAis Gestion Des Ressources Hydriques BbaIslam AissiPas encore d'évaluation
- Analyse de Groupe Et Psychodrame-2002Document317 pagesAnalyse de Groupe Et Psychodrame-2002Mallis FatiPas encore d'évaluation
- Protocole Originale PowerpointDocument59 pagesProtocole Originale Powerpointعمد ابن الفPas encore d'évaluation
- TD MPC Gmsi MrmiDocument4 pagesTD MPC Gmsi MrmiMohammed El azhariPas encore d'évaluation
- Nefoussi Hafsa Memoire Master 02Document78 pagesNefoussi Hafsa Memoire Master 02Rafik Armadilo100% (1)
- CH 1 AtomeDocument7 pagesCH 1 AtomeAnish SundararajanPas encore d'évaluation