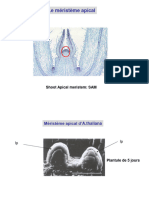
Vous aimerez peut-être aussi
- Les 72 Génies de KabbaleDocument133 pagesLes 72 Génies de KabbaleFernando Rodrigues Lopes100% (3)
- Neurobiologie: Les Grands Articles d'UniversalisD'EverandNeurobiologie: Les Grands Articles d'UniversalisPas encore d'évaluation
- Pestel ReferencesDocument108 pagesPestel ReferencesMed MomoPas encore d'évaluation
- Supplement CarieDocument24 pagesSupplement Carielamia temmouchePas encore d'évaluation
- BIOLOGIE Terminales SE SS SMDocument102 pagesBIOLOGIE Terminales SE SS SMOusmane Sylla100% (2)
- Gazoline Juin 2017Document148 pagesGazoline Juin 2017Mou Mou100% (2)
- TD Génétique Des PopulationsDocument10 pagesTD Génétique Des Populationscoursenligne.lille1100% (17)
- La désintoxication douce: Programme de détoxification naturelleD'EverandLa désintoxication douce: Programme de détoxification naturelleÉvaluation : 5 sur 5 étoiles5/5 (1)
- M Ethodes Num Eriques Pour Les Ecoulements IncompressiblesDocument80 pagesM Ethodes Num Eriques Pour Les Ecoulements IncompressiblesMohamed LatifiPas encore d'évaluation
- FFB - Guide Des Déchets DangereuxDocument16 pagesFFB - Guide Des Déchets DangereuxProutPas encore d'évaluation
- Al7sn02tdpa0112 Sequence 04Document104 pagesAl7sn02tdpa0112 Sequence 04A100% (1)
- H50D H60D H70D H80D de 01.2004 LINDEDocument104 pagesH50D H60D H70D H80D de 01.2004 LINDEAmmar ZouhairPas encore d'évaluation
- Exos Genetique Des PopDocument92 pagesExos Genetique Des PopFarrouha Haddad100% (1)
- 001700doc04[1]Document38 pages001700doc04[1]hanane boutajPas encore d'évaluation
- Quelle Est L'origine de La Variation de Colorations Des Rayures Chez Les Poissons-Zèbres ?Document1 pageQuelle Est L'origine de La Variation de Colorations Des Rayures Chez Les Poissons-Zèbres ?Louka CharlierPas encore d'évaluation
- Support Cours Agad L3 PNA 2223 (CHAPITRE 01) 1 Sect 01 070123Document6 pagesSupport Cours Agad L3 PNA 2223 (CHAPITRE 01) 1 Sect 01 070123benhammanaPas encore d'évaluation
- COURS DE GENETIQUE MOLECUlAIREDocument78 pagesCOURS DE GENETIQUE MOLECUlAIREDaim OndsPas encore d'évaluation
- Devoir Bilan Chap Biodiversite T05 Nov 23Document4 pagesDevoir Bilan Chap Biodiversite T05 Nov 23Chouaib BouhouchPas encore d'évaluation
- Chapitres 3 & 4Document17 pagesChapitres 3 & 4Njiasse MaazouPas encore d'évaluation
- Support TD Génétique - S5 - 2017 - 2018 - PR Achtak PDFDocument8 pagesSupport TD Génétique - S5 - 2017 - 2018 - PR Achtak PDFZineb Dahmani50% (2)
- Chapitre III PDFDocument9 pagesChapitre III PDFyounes chouaiPas encore d'évaluation
- 04fc7f - (Full Permission) PDFDocument3 pages04fc7f - (Full Permission) PDFLotfi TouilPas encore d'évaluation
- La Biodiversité Et Son Évolution Thème 3 - Chapitre 1: RappelDocument6 pagesLa Biodiversité Et Son Évolution Thème 3 - Chapitre 1: Rappelzp98h7jbcwPas encore d'évaluation
- Série D'exercices: Exercice 1Document3 pagesSérie D'exercices: Exercice 1fardadi rachidPas encore d'évaluation
- Biolo Tse p1Document14 pagesBiolo Tse p1Sawoulou celest DopavoguiPas encore d'évaluation
- TD GP 2018 HWDocument4 pagesTD GP 2018 HWFati ZoraPas encore d'évaluation
- TD Gen Pop Agro2Document6 pagesTD Gen Pop Agro2Freenel BossaPas encore d'évaluation
- Vrai TD de Génétique 2019Document5 pagesVrai TD de Génétique 2019Moussa KonéPas encore d'évaluation
- L2 S4 BOP Cuguen Genetique PDFDocument149 pagesL2 S4 BOP Cuguen Genetique PDFFarrouha Haddad0% (1)
- tes-t3-ci-doc-ele-cours-chapitreDocument13 pagestes-t3-ci-doc-ele-cours-chapitrehicham IdboufkerPas encore d'évaluation
- TD - 1 - L3 - Genomique - PPTX - Filename - UTF-8''TD (1) L3 GenomiqueDocument31 pagesTD - 1 - L3 - Genomique - PPTX - Filename - UTF-8''TD (1) L3 GenomiqueMĄd Loubna TebibPas encore d'évaluation
- La CGHDocument5 pagesLa CGHdyhp2pswsrPas encore d'évaluation
- Fiches Noel TerminaleDocument10 pagesFiches Noel Terminalekeimone NZABAPas encore d'évaluation
- SN16-TE-01-19 - Chapitre 3Document8 pagesSN16-TE-01-19 - Chapitre 3Sadek KkkPas encore d'évaluation
- TG ES BiodiversitéDocument46 pagesTG ES BiodiversitéMorgane MarlangePas encore d'évaluation
- 1 - Technique de Biologie Et Génétique Moléculaire-2Document4 pages1 - Technique de Biologie Et Génétique Moléculaire-2Guillaume MORADEIPas encore d'évaluation
- TP 1 Séquençage Et Séparation Des GénomesDocument4 pagesTP 1 Séquençage Et Séparation Des GénomesMaxPas encore d'évaluation
- Evaluation formative 2trim TLEC M F.FAROUD23-24Document8 pagesEvaluation formative 2trim TLEC M F.FAROUD23-24oswald kagbahintoPas encore d'évaluation
- Ia de Diourbel Gscab Premiere S1/S2 Serie Exercices 20123: Exercice 1Document3 pagesIa de Diourbel Gscab Premiere S1/S2 Serie Exercices 20123: Exercice 1KhadyPas encore d'évaluation
- Intro GenomiqueDocument51 pagesIntro Genomiquefatima zohra mahammiPas encore d'évaluation
- Comores Sujet C Science (SVT) Bac 2011Document4 pagesComores Sujet C Science (SVT) Bac 2011Mouhammad Hassani HamadiPas encore d'évaluation
- 11-Organistaion Du Génome HumainDocument38 pages11-Organistaion Du Génome HumainMellifluous LullabyPas encore d'évaluation
- 1) Statistiques DescriptivesDocument9 pages1) Statistiques DescriptivesahlemmPas encore d'évaluation
- Arguments Pour La GenetiqueDocument3 pagesArguments Pour La Genetiquemennarais55Pas encore d'évaluation
- Les Marqueurs MoléculairesDocument3 pagesLes Marqueurs MoléculairesAhmed Younsi100% (1)
- TP TD1 Clones Et MutationsDocument9 pagesTP TD1 Clones Et Mutationsdiopstart11Pas encore d'évaluation
- Cameroun-2017-BAC-SVT-Series-C-TIDocument8 pagesCameroun-2017-BAC-SVT-Series-C-TIeviwounoukingPas encore d'évaluation
- CR 11 - Méthodes D'investigations en Pathologies MoleculairesDocument89 pagesCR 11 - Méthodes D'investigations en Pathologies MoleculairesMouad HiliaPas encore d'évaluation
- 1 - Introduction À La GénomiqueDocument22 pages1 - Introduction À La Génomiquea.itatahine100% (1)
- Terminale 2020 2021 Rappels de 1èreDocument3 pagesTerminale 2020 2021 Rappels de 1èreIman QPas encore d'évaluation
- 4 Session Rattrapage SVT 2017Document11 pages4 Session Rattrapage SVT 2017merzougPas encore d'évaluation
- 1388 Outils de La Biologie Moleculaire M AberkaneDocument86 pages1388 Outils de La Biologie Moleculaire M AberkaneZakariya nadirPas encore d'évaluation
- QCM Avec Key SVTDocument22 pagesQCM Avec Key SVTIzem AtlasPas encore d'évaluation
- ExamenPROBAS 08-01-20 CorrigeDocument5 pagesExamenPROBAS 08-01-20 CorrigeFJ8o ooo.oQUIMPERPas encore d'évaluation
- BAC Blanc 2024 SERIE D SVT Dren Abidjan 3 (Cocody)Document3 pagesBAC Blanc 2024 SERIE D SVT Dren Abidjan 3 (Cocody)46682504.isPas encore d'évaluation
- Sujet Et Corrige Bac Blanc 2023 SVT Serie C Regional de Katiola Cote D'ivoireDocument7 pagesSujet Et Corrige Bac Blanc 2023 SVT Serie C Regional de Katiola Cote D'ivoireJules Marc Romain AchiPas encore d'évaluation
- Protéomique: Approche Nécessaire Physiologie Et Génétique PlantesDocument9 pagesProtéomique: Approche Nécessaire Physiologie Et Génétique Plantesnaomiesolefack03Pas encore d'évaluation
- Divisions Cellulaires ActiviteDocument3 pagesDivisions Cellulaires ActiviteRLXMPas encore d'évaluation
- Les Différents Types D'hybridation MoléculaireDocument10 pagesLes Différents Types D'hybridation Moléculairesherry1808Pas encore d'évaluation
- Correction Ex GDP M1BIPDocument7 pagesCorrection Ex GDP M1BIPabdoul djalil soumanaPas encore d'évaluation
- TP de Génétique TP B11. Outils Pour L'étude Des Génomes: I. Etablir Une Carte de RestrictionDocument10 pagesTP de Génétique TP B11. Outils Pour L'étude Des Génomes: I. Etablir Une Carte de Restrictionmoussodjirebecca9Pas encore d'évaluation
- Genetique TD Corrige 11.docx (0Document8 pagesGenetique TD Corrige 11.docx (0Nathan DjeranePas encore d'évaluation
- Term A Cours Modèles de Harding Term ES Chap1Document5 pagesTerm A Cours Modèles de Harding Term ES Chap1bouangaPas encore d'évaluation
- Hal 00901100Document7 pagesHal 00901100Mama GueyePas encore d'évaluation
- Burkina 2016 SVT Serie D 2nd Tour S2Document4 pagesBurkina 2016 SVT Serie D 2nd Tour S2bigtourePas encore d'évaluation
- PDF document 2Document2 pagesPDF document 2joellesonyaPas encore d'évaluation
- La Biodiversité Et Son Évolution - Tle - Cours en…Document2 pagesLa Biodiversité Et Son Évolution - Tle - Cours en…joellesonyaPas encore d'évaluation
- Sujet L3 BPDP Déc 2014Document15 pagesSujet L3 BPDP Déc 2014sextansPas encore d'évaluation
- L3 1 2014-2015 FinalDocument66 pagesL3 1 2014-2015 FinalsextansPas encore d'évaluation
- Licence SDV Rentrée 2015 L3 CarreireDocument31 pagesLicence SDV Rentrée 2015 L3 CarreiresextansPas encore d'évaluation
- L3 SD Juin 2012Document1 pageL3 SD Juin 2012sextansPas encore d'évaluation
- Cours 4 BDV 2014Document93 pagesCours 4 BDV 2014sextansPas encore d'évaluation
- TD Genetique Populations 2016Document9 pagesTD Genetique Populations 2016sextansPas encore d'évaluation
- Contrat Pédagogique Licence ST 2014 2015Document8 pagesContrat Pédagogique Licence ST 2014 2015sextansPas encore d'évaluation
- OptiqueSVTE Doc Etudiant 2014-15Document55 pagesOptiqueSVTE Doc Etudiant 2014-15sextansPas encore d'évaluation
- Cours 2011 SE1005 5 JLSDocument44 pagesCours 2011 SE1005 5 JLSsextansPas encore d'évaluation
- Cours 2011 SE1005 2 JLSDocument39 pagesCours 2011 SE1005 2 JLSsextansPas encore d'évaluation
- Cours 2011 SE1005 6 JLSDocument16 pagesCours 2011 SE1005 6 JLSsextansPas encore d'évaluation
- Cours 2011 SE1005 8 JLSDocument42 pagesCours 2011 SE1005 8 JLSsextansPas encore d'évaluation
- Calendrier Licence 2015-2016Document2 pagesCalendrier Licence 2015-2016sextansPas encore d'évaluation
- Chap1 SeriecDocument33 pagesChap1 SeriecsextansPas encore d'évaluation
- Resume Cours 3Document1 pageResume Cours 3Thomas HansenPas encore d'évaluation
- Chapitre 5: Les Interactions Intermoléculaires/ Etats de La MatièreDocument16 pagesChapitre 5: Les Interactions Intermoléculaires/ Etats de La MatièresextansPas encore d'évaluation
- Cours 2011 SE1005 3 JLSDocument15 pagesCours 2011 SE1005 3 JLSsextansPas encore d'évaluation
- Resume Cours 9Document1 pageResume Cours 9sextansPas encore d'évaluation
- DéfinitionsDocument3 pagesDéfinitionssextansPas encore d'évaluation
- Cours 9 - Reproduction - Sexuee - Angiospermes - Graines-Fruit-StrategiesDocument22 pagesCours 9 - Reproduction - Sexuee - Angiospermes - Graines-Fruit-StrategiessextansPas encore d'évaluation
- Chap2 Diversite Vivant 2012Document73 pagesChap2 Diversite Vivant 2012Thomas HansenPas encore d'évaluation
- Diapos Cours Olfaction Fenelon 2015 Universtie de BordeauxDocument72 pagesDiapos Cours Olfaction Fenelon 2015 Universtie de BordeauxsextansPas encore d'évaluation
- Chapitre 1-Domaine D'étude de La MicrobiologieDocument100 pagesChapitre 1-Domaine D'étude de La MicrobiologieBernard SoumaPas encore d'évaluation
- ISO 11133 (1) (Mode de Compatibilité)Document9 pagesISO 11133 (1) (Mode de Compatibilité)agrojus778790Pas encore d'évaluation
- Nouvelles RecettesDocument27 pagesNouvelles RecettesSimon GrégoirePas encore d'évaluation
- Chapitre IDocument15 pagesChapitre Ihassane ayiPas encore d'évaluation
- PR ésentation 2Document17 pagesPR ésentation 2oumqsdfPas encore d'évaluation
- CNTRL 2 Bac SC X S 1 - 22-23Document2 pagesCNTRL 2 Bac SC X S 1 - 22-23Outouli MohamedPas encore d'évaluation
- Cours Complet de TopologieDocument60 pagesCours Complet de TopologieKygoPas encore d'évaluation
- SLUMP TEST+ COMPRESSION - NidhiDocument6 pagesSLUMP TEST+ COMPRESSION - NidhiHrithik IGPas encore d'évaluation
- Cours Système NerveuxDocument9 pagesCours Système NerveuxMohammed errifaiyPas encore d'évaluation
- Book - Présentation de La Société MGT Apporteur de Solution MécaniqueDocument44 pagesBook - Présentation de La Société MGT Apporteur de Solution MécaniqueASMGTPas encore d'évaluation
- Seance Du 05-06-2020Document2 pagesSeance Du 05-06-2020Chimie PhysiquePas encore d'évaluation
- La Santé Mentale PrésentationDocument12 pagesLa Santé Mentale PrésentationFerdaous MalamanePas encore d'évaluation
- Jeu Du Bingo Interctif ANIMAUXDocument14 pagesJeu Du Bingo Interctif ANIMAUXElvira QuliyevaPas encore d'évaluation
- TopoDocument3 pagesTopoSta ToxPas encore d'évaluation
- Chapitre 12 StatistiquesDocument25 pagesChapitre 12 StatistiquesLyse NdifoPas encore d'évaluation
- La Ministre de La Justice Est Responsable de L'application de La Présente Loi. Décret 919-2019 Du 4 Septembre 2019, (2019) 151 G.O. 2, 3975Document10 pagesLa Ministre de La Justice Est Responsable de L'application de La Présente Loi. Décret 919-2019 Du 4 Septembre 2019, (2019) 151 G.O. 2, 3975David MessierPas encore d'évaluation
- PDFDocument267 pagesPDFomariPas encore d'évaluation
- Mecanique Tp3usmba09Document8 pagesMecanique Tp3usmba09AymanPas encore d'évaluation
- Roxtec Ex Product Catalogue en FR de Es ItDocument88 pagesRoxtec Ex Product Catalogue en FR de Es Itmonik_atabaresPas encore d'évaluation
- 1741 DGD 4Document2 pages1741 DGD 4SerignePas encore d'évaluation
- 354498532-Not E-De-Cal Cul-V RD - Wat ErmarkDocument8 pages354498532-Not E-De-Cal Cul-V RD - Wat ErmarkIbrahima Laye BarrePas encore d'évaluation
- Caddy Dec2022 PDFDocument12 pagesCaddy Dec2022 PDFHabanos CubanoPas encore d'évaluation
- TP Api IiDocument11 pagesTP Api IiAdem AounPas encore d'évaluation













![001700doc04[1]](https://imgv2-2-f.scribdassets.com/img/document/723614130/149x198/022376341a/1713279802?v=1)