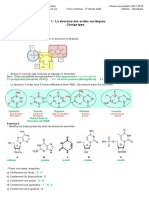
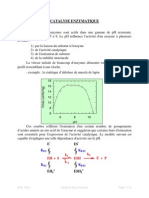
Vous aimerez peut-être aussi
- Les Principaux Défauts de SoudureDocument7 pagesLes Principaux Défauts de SoudureAli Clubist100% (3)
- Cours 2 - Notions de PHDocument3 pagesCours 2 - Notions de PHwebmastercours100% (4)
- Acides nucléiques: Les Grands Articles d'UniversalisD'EverandAcides nucléiques: Les Grands Articles d'UniversalisPas encore d'évaluation
- Les Additifs AlimentairesDocument70 pagesLes Additifs AlimentairesAnass DraPas encore d'évaluation
- Biologie Moleculaire - Chap1-Structure Des Acides NucleiquesDocument8 pagesBiologie Moleculaire - Chap1-Structure Des Acides NucleiquesMustapha SamiPas encore d'évaluation
- Projet Gitologie SurDocument12 pagesProjet Gitologie SurĂlaa ÊđinePas encore d'évaluation
- Base de L'hereditéDocument67 pagesBase de L'hereditéNanitta ChhadePas encore d'évaluation
- Epreuve de Biochimie PDFDocument6 pagesEpreuve de Biochimie PDFZoulkiffiPas encore d'évaluation
- Résumé Essais de DuretéDocument4 pagesRésumé Essais de Duretéihebchaieb0% (1)
- Generalité Sur ADNDocument4 pagesGeneralité Sur ADNMedecine DentairePas encore d'évaluation
- Genetique1an16 AdnDocument11 pagesGenetique1an16 AdnhichamPas encore d'évaluation
- I - L'Adn 1. Les Constituants de l'ADNDocument13 pagesI - L'Adn 1. Les Constituants de l'ADNZakaria FerkachePas encore d'évaluation
- Chapter I - Structures Des Acide Nucleiques - 2020Document25 pagesChapter I - Structures Des Acide Nucleiques - 2020julien KoudouPas encore d'évaluation
- Génétique MoléculaireDocument14 pagesGénétique MoléculaireBelhadj BahiaPas encore d'évaluation
- Structure Des Acides NucleiquesDocument9 pagesStructure Des Acides NucleiquesagbodrepierrettePas encore d'évaluation
- Cours - 02 - Nature Chimiques Du Matériel GénétiqueDocument34 pagesCours - 02 - Nature Chimiques Du Matériel Génétiquejames deyPas encore d'évaluation
- 1 Acides NuclDocument7 pages1 Acides Nuclhelmi abbessiPas encore d'évaluation
- Biologie Moleculaire MunirDocument42 pagesBiologie Moleculaire MunirOuu SsaamaPas encore d'évaluation
- Biologie FondamentaleDocument5 pagesBiologie FondamentaleainaneayoubPas encore d'évaluation
- An EstDocument33 pagesAn EstHamza AzouitinPas encore d'évaluation
- Cours BiomolDocument144 pagesCours BiomolHajar ChehmatPas encore d'évaluation
- 2 - Structure Et Propriété Des Acides NucléiquesDocument9 pages2 - Structure Et Propriété Des Acides NucléiquesEric OuakaraPas encore d'évaluation
- Cours - 02 - Nature Chimiques Du Matériel GénétiqueDocument31 pagesCours - 02 - Nature Chimiques Du Matériel GénétiqueMohamed Akli TigrinePas encore d'évaluation
- Acides NucléiquesDocument23 pagesAcides NucléiquesKEBIR ZAHRAPas encore d'évaluation
- Le Génie Génétique AgroDocument112 pagesLe Génie Génétique AgroWa LidPas encore d'évaluation
- Biochimie1an Poly-Structure Proprietes Acides Nucleiques2020feragaDocument12 pagesBiochimie1an Poly-Structure Proprietes Acides Nucleiques2020feragamalak zerizerPas encore d'évaluation
- Cours BioMolDocument209 pagesCours BioMolayasannahi123Pas encore d'évaluation
- Sequence 6 - Les Acides NucleiquesDocument85 pagesSequence 6 - Les Acides Nucleiquesfijanu4Pas encore d'évaluation
- ADN. StructureDocument9 pagesADN. Structuredark kristalPas encore d'évaluation
- Bio MOL 2020 CoursDocument154 pagesBio MOL 2020 CoursWilkendy BarthélemyPas encore d'évaluation
- Le Support de L'information Génétique l'ADNDocument29 pagesLe Support de L'information Génétique l'ADNMbaye AbouPas encore d'évaluation
- ADN Et ARNDocument48 pagesADN Et ARNantaafall464Pas encore d'évaluation
- Cours Biologie Moleculaire ACQM 2022Document95 pagesCours Biologie Moleculaire ACQM 2022elmajdoubreda11Pas encore d'évaluation
- Biologie MoléculaireDocument12 pagesBiologie MoléculaireDaniela RusuPas encore d'évaluation
- ADN BCM1501 2021 - Part 1Document62 pagesADN BCM1501 2021 - Part 1supernovaPas encore d'évaluation
- Les Acides Nucleiques 20232024Document7 pagesLes Acides Nucleiques 20232024rla320331Pas encore d'évaluation
- Reproduction Du Génotype Au PhénotypeDocument7 pagesReproduction Du Génotype Au Phénotypeorion.13ferrPas encore d'évaluation
- Séance 1 Acides Nucléiques Et Information GénétiqueDocument68 pagesSéance 1 Acides Nucléiques Et Information GénétiqueMbaye AbouPas encore d'évaluation
- Chap 1 ADNDocument26 pagesChap 1 ADNHassanPas encore d'évaluation
- MIB 3086 Chapitre 1 Et 2Document17 pagesMIB 3086 Chapitre 1 Et 2BOULAMA HASSANA MASSAWPas encore d'évaluation
- Biologie Cours 1Document7 pagesBiologie Cours 1syedsakeenahafafPas encore d'évaluation
- Acides Nucléiques 2020 - 2021Document33 pagesAcides Nucléiques 2020 - 2021Rottina RossyPas encore d'évaluation
- Atelier Bioinfo S1Document53 pagesAtelier Bioinfo S1Lamili AyoubPas encore d'évaluation
- Biochimie 07 - Les Acides Nucéiques Et Structure Du GénomeDocument26 pagesBiochimie 07 - Les Acides Nucéiques Et Structure Du GénomeNathan CohenPas encore d'évaluation
- Cours Des Acides NucleiquesDocument63 pagesCours Des Acides Nucleiquesfouad abdelhamid100% (2)
- LADNDocument14 pagesLADNthamielou01Pas encore d'évaluation
- 11.02.2016 Les ProtéinesDocument29 pages11.02.2016 Les ProtéinesEaPas encore d'évaluation
- ABCCH1Document41 pagesABCCH1Sambardé FstPas encore d'évaluation
- Les ARN PDFDocument6 pagesLes ARN PDFMarion MilletPas encore d'évaluation
- TD Biologie Moléculaire BPR 2021Document21 pagesTD Biologie Moléculaire BPR 2021rafaramalala1307Pas encore d'évaluation
- Cours Adn Et Arn Et Linformation Genetique Master 1Document13 pagesCours Adn Et Arn Et Linformation Genetique Master 1t8zlhqcvtaciizli8qwPas encore d'évaluation
- Cours1 BMGGDocument5 pagesCours1 BMGGsalama2011Pas encore d'évaluation
- TD1 QCMDocument13 pagesTD1 QCMjebranesanaa92Pas encore d'évaluation
- Chapitre 2-Notions Fondamentales de Biologie Et de GénétiqueDocument46 pagesChapitre 2-Notions Fondamentales de Biologie Et de GénétiqueDiop MakerPas encore d'évaluation
- SynthèseDocument10 pagesSynthèseElvy prices Sampinou mpompaPas encore d'évaluation
- 3.1 Structure Et Propriétés Des Nucléotides Et Acis NucléiquesDocument39 pages3.1 Structure Et Propriétés Des Nucléotides Et Acis NucléiquessedighdahmanPas encore d'évaluation
- Cours 4 - DIAPO - ADN Aux ProtéinesDocument40 pagesCours 4 - DIAPO - ADN Aux Protéinesbyy.monkey emzPas encore d'évaluation
- 1-Structure Bases, Nucléotides Et Acides Nucléiques ARN/ADNDocument8 pages1-Structure Bases, Nucléotides Et Acides Nucléiques ARN/ADNZineb DahmaniPas encore d'évaluation
- TraductionDocument36 pagesTraductionnonamethe427Pas encore d'évaluation
- Acides Nucleiques Cours 01Document41 pagesAcides Nucleiques Cours 01MAITRE ValeryPas encore d'évaluation
- Dossier OcDocument10 pagesDossier OcEva BermudezPas encore d'évaluation
- Tweet: La Synthèse Des ProtéinesDocument59 pagesTweet: La Synthèse Des ProtéinesNadaPas encore d'évaluation
- UE1 Biochimie 6 - Structure Du GénomeDocument18 pagesUE1 Biochimie 6 - Structure Du Génomesun-nee-chan9Pas encore d'évaluation
- Présentation (2022) - Nucléotides Et Acides NucléiquesDocument68 pagesPrésentation (2022) - Nucléotides Et Acides Nucléiquesmintchimechak2Pas encore d'évaluation
- Corrigé Type Du TD1 Génétique (La Structure Des Acides Nucléiques)Document3 pagesCorrigé Type Du TD1 Génétique (La Structure Des Acides Nucléiques)Doudou Amine50% (2)
- 5 EstomacDocument6 pages5 Estomacsevda mouviePas encore d'évaluation
- 3-Physiologie GastriqueDocument5 pages3-Physiologie Gastriquesevda mouviePas encore d'évaluation
- 5-Digestion Et Absorption IntestinaleDocument3 pages5-Digestion Et Absorption Intestinalesevda mouviePas encore d'évaluation
- 13-Outils de Biologie Moléculaire 1 N.MDocument5 pages13-Outils de Biologie Moléculaire 1 N.Msevda mouviePas encore d'évaluation
- 10-Modifications Post Traductionnelle N.MDocument3 pages10-Modifications Post Traductionnelle N.Msevda mouviePas encore d'évaluation
- Communication CellulaireDocument4 pagesCommunication Cellulairesevda mouviePas encore d'évaluation
- Métabolisme Des LipidesDocument7 pagesMétabolisme Des Lipidessevda mouviePas encore d'évaluation
- Med 1an Emd2 Info16Document6 pagesMed 1an Emd2 Info16sevda mouviePas encore d'évaluation
- Dosage D Un Anti Mousse 1Document2 pagesDosage D Un Anti Mousse 1غرباءPas encore d'évaluation
- La Transformation Du Petrole Et Ses Derives Vincent DeshayeDocument9 pagesLa Transformation Du Petrole Et Ses Derives Vincent DeshayeKadi MiragePas encore d'évaluation
- AT - Se Paration de Prote Ines Par SDS PAGEDocument8 pagesAT - Se Paration de Prote Ines Par SDS PAGErahmani100% (1)
- 4 - Catalyse Enzymatique PDFDocument14 pages4 - Catalyse Enzymatique PDFPrince Abdoo100% (1)
- 9852 2043 03b Maintenance Instructions ROC L6 (25), L6 (30), L8 (25), L8Document124 pages9852 2043 03b Maintenance Instructions ROC L6 (25), L6 (30), L8 (25), L8agvassPas encore d'évaluation
- PARTIE 1. Biochimie Métabolique (Enregistré Automatiquement) - 1Document42 pagesPARTIE 1. Biochimie Métabolique (Enregistré Automatiquement) - 1beauty queensPas encore d'évaluation
- Complexe TDDocument4 pagesComplexe TDHamza SidhmPas encore d'évaluation
- Série D'exercices - Les Alcanes - 1e S - SunudaaraDocument7 pagesSérie D'exercices - Les Alcanes - 1e S - Sunudaarapddj4t2w58Pas encore d'évaluation
- Des Définitions: TP N1: Préparation D'une Solution D'acide Faible, L'acide OxaliqueDocument3 pagesDes Définitions: TP N1: Préparation D'une Solution D'acide Faible, L'acide Oxaliquesidi mohamed el amine nekkalPas encore d'évaluation
- TleD - CH - L12 - Acides Alpha AminésDocument7 pagesTleD - CH - L12 - Acides Alpha AminésHamadi DialloPas encore d'évaluation
- Contrôle de L'eau À Usage PharmaceutiqueDocument88 pagesContrôle de L'eau À Usage PharmaceutiqueRochdi KherratPas encore d'évaluation
- CRF M.M A Retenir en Physique Et ChimieDocument7 pagesCRF M.M A Retenir en Physique Et ChimieIdrissa KouyatéPas encore d'évaluation
- PDF QCM Materiaux - CorrigeDocument1 pagePDF QCM Materiaux - Corrigekzi ita100% (4)
- 1ers Chap6 TP10Document3 pages1ers Chap6 TP10Nadia Ait AhmedPas encore d'évaluation
- Fiche Technique - Joints Toriques EP851Document1 pageFiche Technique - Joints Toriques EP851Nicolas AguilarPas encore d'évaluation
- CombinepdfDocument96 pagesCombinepdfHALIMO FarikoPas encore d'évaluation
- Cours PCT A TIRERDocument60 pagesCours PCT A TIRERJoy TowendoPas encore d'évaluation
- tp05 Maiver 2Document14 pagestp05 Maiver 2kmeriemPas encore d'évaluation
- Polissage Électrolytique Du CuivreDocument6 pagesPolissage Électrolytique Du Cuivrekhouloud amaraPas encore d'évaluation
- Guide Enderbot Apres Maj 2019 by TeegrewDocument9 pagesGuide Enderbot Apres Maj 2019 by TeegrewKarinr HdhdhfPas encore d'évaluation
- Guide Environnemental Sur L'ApplicationdespeinturesDocument54 pagesGuide Environnemental Sur L'ApplicationdespeinturesyassinPas encore d'évaluation
- CyanuresDocument5 pagesCyanuresyou tubiaPas encore d'évaluation
- Penguard Express: Fiche TechniqueDocument5 pagesPenguard Express: Fiche Techniqueali piPas encore d'évaluation
- Iutmosel Polymeres 2012 CHDocument2 pagesIutmosel Polymeres 2012 CHAbdallah AmmarPas encore d'évaluation
- Fuel Recovery From Waste Oily Sludge Using Solvent ExtractionDocument3 pagesFuel Recovery From Waste Oily Sludge Using Solvent ExtractionKinda DolaPas encore d'évaluation