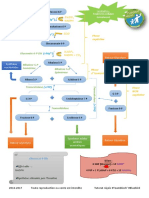
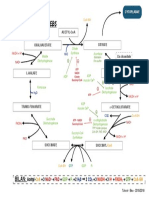
Vous aimerez peut-être aussi
- Applications de la spectrophotomérie en phytochimie: sciencesD'EverandApplications de la spectrophotomérie en phytochimie: sciencesPas encore d'évaluation
- Cycle de KrebsDocument37 pagesCycle de KrebscepPas encore d'évaluation
- C. Biochimie III. Lipides (III.1. Métabolisme Des Acides Gras)Document41 pagesC. Biochimie III. Lipides (III.1. Métabolisme Des Acides Gras)Rania HamdiPas encore d'évaluation
- CHAP7 MET DES LIPIDES 1 Métabolisme Des Acides Gras 2022 2023 PLUSDocument9 pagesCHAP7 MET DES LIPIDES 1 Métabolisme Des Acides Gras 2022 2023 PLUSdianaPas encore d'évaluation
- Biosynthése Des Lipidsynt10Document9 pagesBiosynthése Des Lipidsynt10mhichePas encore d'évaluation
- 1 4 Metabolisme Des Lipides PDFDocument22 pages1 4 Metabolisme Des Lipides PDFAya Bouchareb100% (1)
- Métabolisme Des LipidesDocument47 pagesMétabolisme Des LipidesBoubaker AmiraPas encore d'évaluation
- Métabolisme Des LipidesDocument17 pagesMétabolisme Des LipidesMei Wan100% (1)
- CH3.Lipogénèse PFDocument25 pagesCH3.Lipogénèse PFkhammar hichemPas encore d'évaluation
- Metabolisme Des Acides Gras 2018 PDFDocument85 pagesMetabolisme Des Acides Gras 2018 PDFPace MakerPas encore d'évaluation
- Cycle de Krebs PDFDocument9 pagesCycle de Krebs PDFMa RyPas encore d'évaluation
- Metabo Partie2Document6 pagesMetabo Partie2la biochimiste100% (2)
- Métabolisme Des Acides AminésDocument87 pagesMétabolisme Des Acides AminésBlueskyindan100% (1)
- Bioch2an27 NeoglucogeneseDocument9 pagesBioch2an27 NeoglucogeneseIngenieur AgroPas encore d'évaluation
- Cours - Biochimie Structurale - Lipides - CAG - UNA - 2019 - CopieDocument83 pagesCours - Biochimie Structurale - Lipides - CAG - UNA - 2019 - CopieStellaPas encore d'évaluation
- Metabolismes Des Glucides PDFDocument151 pagesMetabolismes Des Glucides PDFMa RyPas encore d'évaluation
- Chap 1 Les Glucides Structure Et MétabolismeDocument75 pagesChap 1 Les Glucides Structure Et Métabolismeafaf gou100% (1)
- 12-Les CatecholaminesDocument4 pages12-Les CatecholaminesoutsiderPas encore d'évaluation
- Métabolisme LipidiqueDocument2 pagesMétabolisme LipidiqueAro Manda100% (1)
- Voie Des Pentoses Phosphates (MaJ) - 2Document1 pageVoie Des Pentoses Phosphates (MaJ) - 2Hadrien De Greef100% (2)
- 5 Metabolisme 10 11Document23 pages5 Metabolisme 10 11Slip Horticole100% (1)
- NeoglucogeneseDocument35 pagesNeoglucogenesecepPas encore d'évaluation
- Le Cycle de KrebsDocument19 pagesLe Cycle de KrebsLandry RimbaybarPas encore d'évaluation
- Correction TD Metabolisme Glucidique 2Document31 pagesCorrection TD Metabolisme Glucidique 2acheparfaite958Pas encore d'évaluation
- Chapitre Voies MétaboliquesDocument57 pagesChapitre Voies MétaboliquesMBAÏADOUM NGARGUINAM RodriguePas encore d'évaluation
- TD 2 de Biochimie StructuraleDocument10 pagesTD 2 de Biochimie Structuraleachouake100% (1)
- Métabolisme Des Glucides Fiche PDFDocument7 pagesMétabolisme Des Glucides Fiche PDFNaïs LABBADIPas encore d'évaluation
- Metabolisme Des TriglycéridesDocument24 pagesMetabolisme Des Triglycéridesvague2000Pas encore d'évaluation
- Métabolisme Des GlucidesDocument28 pagesMétabolisme Des GlucidesilaPas encore d'évaluation
- 2 - TD Métabolisme LipidiqueDocument4 pages2 - TD Métabolisme LipidiqueJina ThiPas encore d'évaluation
- TD 3 Corrigé Regulation SABRINA.2019Document5 pagesTD 3 Corrigé Regulation SABRINA.2019axelPas encore d'évaluation
- TD Acides Aminés Et PeptidesDocument2 pagesTD Acides Aminés Et PeptidesNaruto & SasukePas encore d'évaluation
- Correction TDDocument21 pagesCorrection TDKarim Pichou0% (1)
- Enzymologie PDFDocument28 pagesEnzymologie PDFTCHEMMIK NazihaPas encore d'évaluation
- S1 TD2 Protides PDFDocument8 pagesS1 TD2 Protides PDFmartinmune67% (3)
- 02 Structure Cyclique Des OsesDocument10 pages02 Structure Cyclique Des OsesBook Hunter100% (1)
- Bioch Corrigé de La Série de TD Sur Les LipidesDocument2 pagesBioch Corrigé de La Série de TD Sur Les LipidesAbdelghani Fellague chebraPas encore d'évaluation
- 2023 - Examen de Biochimie StructuraleDocument2 pages2023 - Examen de Biochimie StructuraleAmadou Konfe100% (1)
- Cinétique EnzymatiqueDocument3 pagesCinétique EnzymatiqueChaka TraorePas encore d'évaluation
- ENZYMOLOGIEDocument17 pagesENZYMOLOGIELoulou LoulaPas encore d'évaluation
- TD QCM Oct 2012Document5 pagesTD QCM Oct 2012Sabri CherrakPas encore d'évaluation
- CM Regulation Metabolique 2 Enzymes Et Allosterie 2019 ModDocument31 pagesCM Regulation Metabolique 2 Enzymes Et Allosterie 2019 ModSirina LadjPas encore d'évaluation
- Biotransformation PDFDocument22 pagesBiotransformation PDFLinaMimosaPas encore d'évaluation
- Les Acides AminesDocument7 pagesLes Acides Aminesbouguidima_sanaPas encore d'évaluation
- TD Protéines (Énoncés)Document4 pagesTD Protéines (Énoncés)Eslem Islam100% (2)
- ExerciceDocument5 pagesExerciceOthmen Bachaa67% (3)
- Chap.3 ProtéineDocument46 pagesChap.3 ProtéineIngenieur AgroPas encore d'évaluation
- 1 1-Introduction PDFDocument21 pages1 1-Introduction PDFحامدي يعقوبPas encore d'évaluation
- Cours OsesDocument76 pagesCours OsesNabil holmes50% (2)
- Biochimie MetaboliqueDocument360 pagesBiochimie MetaboliqueMafankoloPas encore d'évaluation
- TD, Aa, Prot, AnDocument6 pagesTD, Aa, Prot, Ansarahmboumba803Pas encore d'évaluation
- La NéoglucogenèseDocument34 pagesLa Néoglucogenèsevague2000Pas encore d'évaluation
- 1 Glucides Medecine 1 1Document43 pages1 Glucides Medecine 1 1Ali SegueniPas encore d'évaluation
- Acides Aminés Pour Aller Plus LoinDocument15 pagesAcides Aminés Pour Aller Plus LoinCooper WaynePas encore d'évaluation
- Cours Lipides - Part 1Document74 pagesCours Lipides - Part 1AhmedPas encore d'évaluation
- Corrigé de L'examen de Remplacement de Biochimie (2016)Document7 pagesCorrigé de L'examen de Remplacement de Biochimie (2016)elhidhab2014Pas encore d'évaluation
- Guide GlycogénoseDocument73 pagesGuide GlycogénoseVeronique HamelPas encore d'évaluation
- Métabolisme Des Lipides I Cours SViS6Document45 pagesMétabolisme Des Lipides I Cours SViS6laila hammouPas encore d'évaluation
- Métabolisme Énergétique 2 LipidesDocument28 pagesMétabolisme Énergétique 2 LipidesAyella ÔëPas encore d'évaluation
- Cycle de KrebsDocument1 pageCycle de KrebsHadrien De GreefPas encore d'évaluation
- Corrigé Série Lipides 2020-2021Document5 pagesCorrigé Série Lipides 2020-2021Dav MotjolopanePas encore d'évaluation
- Composes FonctionnelsDocument17 pagesComposes FonctionnelsPhytomenadione La Vitamine KPas encore d'évaluation
- Corrigé TD3Document7 pagesCorrigé TD3Mariam Ben AbdelouahabPas encore d'évaluation
- Klubprepa Extrait 7517Document10 pagesKlubprepa Extrait 7517HamZa RaouguiPas encore d'évaluation
- 11-Structure Et Fonction Des ACIDES AMINESDocument55 pages11-Structure Et Fonction Des ACIDES AMINESBook Hunter100% (2)
- Sample Chemistry Entrance Exam 2020 - 2021-sDocument7 pagesSample Chemistry Entrance Exam 2020 - 2021-sAli RammalPas encore d'évaluation
- Tpe Chimie OrganiqueDocument27 pagesTpe Chimie Organiquealiabba0002Pas encore d'évaluation
- Nomenclature Des Carbocycles Et Des Hetero CyclesDocument5 pagesNomenclature Des Carbocycles Et Des Hetero CyclesJean-François Abena100% (3)
- Chim 306 - Notes de CoursDocument43 pagesChim 306 - Notes de Coursnatural recipePas encore d'évaluation
- Orga 9Document14 pagesOrga 9SalahPas encore d'évaluation
- Nomenclature Des Composés OrganiquesDocument16 pagesNomenclature Des Composés OrganiquesRuroniPas encore d'évaluation
- Liste Des Medicaments en AlgerieDocument9 pagesListe Des Medicaments en AlgerieAbdeau1Pas encore d'évaluation
- LIPIDES Lipides Saponifiables SimplesDocument71 pagesLIPIDES Lipides Saponifiables SimplesNalesso Mamadou FofanaPas encore d'évaluation
- Leçons SavonDocument10 pagesLeçons SavonBoubacar Balde100% (2)
- Exercices Ac Carboxyliques-DerivesDocument5 pagesExercices Ac Carboxyliques-DerivesJean-François Abena100% (3)
- 1 3 1 2-Exos-esters-AGDocument1 page1 3 1 2-Exos-esters-AGJacques AdjoyiPas encore d'évaluation
- Chap 2Document4 pagesChap 2Ouehnia tynaPas encore d'évaluation
- Cours Bioch Métab Version CP 2 Agro IAV Partie 2Document135 pagesCours Bioch Métab Version CP 2 Agro IAV Partie 2Your fav Aya100% (1)
- Corrigé Type T12023 PDFDocument3 pagesCorrigé Type T12023 PDFSoumia BouguPas encore d'évaluation
- Corrigé Série Lipides 2020-2021Document5 pagesCorrigé Série Lipides 2020-2021Dav Motjolopane100% (1)
- Cours Chimie TsDocument0 pageCours Chimie TsGueda KanePas encore d'évaluation
- Tpe Chimie OrganiqueDocument13 pagesTpe Chimie Organiquefirmin MfououPas encore d'évaluation
- Acide DicarboxyliqueDocument6 pagesAcide DicarboxyliqueFrederic WustPas encore d'évaluation
- TD Chimie Org Corrigé SuiteDocument20 pagesTD Chimie Org Corrigé SuiteBöris LetökössPas encore d'évaluation
- Les Etapes de La GlycolyseDocument7 pagesLes Etapes de La GlycolyseNaruto & SasukePas encore d'évaluation
- Chap4 OrganiqueDocument19 pagesChap4 OrganiqueMostafa McafsPas encore d'évaluation
- Solution Des Exercices - Les Acides Carboxyliques - Ts - SunudaaraDocument7 pagesSolution Des Exercices - Les Acides Carboxyliques - Ts - SunudaaraJule BaPas encore d'évaluation
- Les LipidesDocument8 pagesLes Lipidesyousraghanem23Pas encore d'évaluation
- Catabolisme Des Aa - Vue Generale - CompressedDocument43 pagesCatabolisme Des Aa - Vue Generale - CompressedRaouf ChikhiPas encore d'évaluation