Vous aimerez peut-être aussi
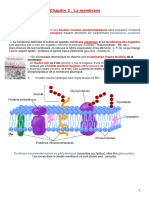
- Chapitre 2. Membrane Plasmique PDFDocument10 pagesChapitre 2. Membrane Plasmique PDFYnés Abidi100% (1)
- Chapitre 2 BiomembraneDocument5 pagesChapitre 2 BiomembraneFatïma ZohraPas encore d'évaluation
- Chapitre II Membrane Plasmique 1Document7 pagesChapitre II Membrane Plasmique 1faresgettasPas encore d'évaluation
- La Memrane PlasmiqueDocument20 pagesLa Memrane PlasmiqueMalik NabazagaPas encore d'évaluation
- (PACES) Physiologie Humaine by Hervé GuénardDocument1 371 pages(PACES) Physiologie Humaine by Hervé Guénardsam egoroffPas encore d'évaluation
- Cours BCG UFR2SDocument19 pagesCours BCG UFR2SSirandou DiaoPas encore d'évaluation
- 02 Revetement-CellulaireDocument6 pages02 Revetement-CellulaireSagno AgnèsPas encore d'évaluation
- Cours Chapitre 2 Membrane PlasmiqueDocument5 pagesCours Chapitre 2 Membrane Plasmiqueayman HammoudaPas encore d'évaluation
- Cours Chapitre 2 Membrane PlasmiqueDocument5 pagesCours Chapitre 2 Membrane PlasmiqueHou TrtyPas encore d'évaluation
- Physiologie de La Cellule Et de La MembraneDocument14 pagesPhysiologie de La Cellule Et de La MembraneKhaoula SahtouriPas encore d'évaluation
- BiomembraneDocument10 pagesBiomembraneSido SidosidoPas encore d'évaluation
- Cours 03. BC (Membrane Plasmique)Document31 pagesCours 03. BC (Membrane Plasmique)KIKOPas encore d'évaluation
- Membranes cellulaires: Les Grands Articles d'UniversalisD'EverandMembranes cellulaires: Les Grands Articles d'UniversalisPas encore d'évaluation
- TD CorrigésDocument19 pagesTD CorrigésPierre Claver Dabou100% (1)
- Chap 2 MPDocument7 pagesChap 2 MPdramohandlhadjPas encore d'évaluation
- La Membrane Plasmique VF CH2Document71 pagesLa Membrane Plasmique VF CH2paul destinPas encore d'évaluation
- Membrane PlasmiqueDocument7 pagesMembrane PlasmiqueSara BoudjemaiPas encore d'évaluation
- Cours 3 - La Membrane PlasmiqueDocument20 pagesCours 3 - La Membrane Plasmiquemanuelakouadio405Pas encore d'évaluation
- Membrane Plasmique - WikipédiaDocument30 pagesMembrane Plasmique - Wikipédiagagnon.jonathan2007Pas encore d'évaluation
- Biologie CellulaireDocument26 pagesBiologie CellulaireBourobou bourobou Chadrack gaelPas encore d'évaluation
- Chapitre1 Biochimie Cellulaire Et FonctionnelleDocument12 pagesChapitre1 Biochimie Cellulaire Et Fonctionnelledraboissouf174Pas encore d'évaluation
- 1.cours Biologie CellulaireDocument63 pages1.cours Biologie Cellulaireبدر الذهابيPas encore d'évaluation
- Biologie Cellulaire: Enseignements DirigesDocument30 pagesBiologie Cellulaire: Enseignements DirigesKourdePas encore d'évaluation
- Le Flux MembranaireDocument22 pagesLe Flux MembranaireYouness KhalfaouiPas encore d'évaluation
- Chapitre 3 La Membrane Plasmique MedDocument6 pagesChapitre 3 La Membrane Plasmique MedDear tynaPas encore d'évaluation
- Biologie Générale HumaineDocument10 pagesBiologie Générale HumaineSarah CabieuPas encore d'évaluation
- Httpsstaff - Univ-Batna2.dzsitesdefaultfilesabdessemed Samirafileschapitre 11 PDFDocument12 pagesHttpsstaff - Univ-Batna2.dzsitesdefaultfilesabdessemed Samirafileschapitre 11 PDFariel.2mariemPas encore d'évaluation
- CHAPITRE. 2 Biomembranes Cour1Document10 pagesCHAPITRE. 2 Biomembranes Cour1Joie De VivrePas encore d'évaluation
- 03 Fonction de La Membrane PlasmiqueDocument9 pages03 Fonction de La Membrane PlasmiqueAbdoul karim SamakePas encore d'évaluation
- Chap 01Document33 pagesChap 01hamidaPas encore d'évaluation
- CH2 - Membrane Plasmique - B.BDocument40 pagesCH2 - Membrane Plasmique - B.Bone asslanPas encore d'évaluation
- Généralité Sur La Membrane PlasmiqueDocument48 pagesGénéralité Sur La Membrane PlasmiqueOussama IdrissPas encore d'évaluation
- Membrane PlasmiqueDocument8 pagesMembrane PlasmiqueGaston CarinePas encore d'évaluation
- 3-Membrane PlasmiqueDocument11 pages3-Membrane Plasmiquekhoualdia marwaPas encore d'évaluation
- 1-Structure Et Archirecture de La Membrane Plasmique NeuronaleDocument8 pages1-Structure Et Archirecture de La Membrane Plasmique NeuronalebayaPas encore d'évaluation
- Membrane Plasmique CH2 2022-2023Document23 pagesMembrane Plasmique CH2 2022-2023Mbaye AbouPas encore d'évaluation
- Cours de Cytologie, Embryologie Et Genetique Medicale by MBAILASSEM MBAINAIDA GEORGESDocument102 pagesCours de Cytologie, Embryologie Et Genetique Medicale by MBAILASSEM MBAINAIDA GEORGESMãhąmãţ Hãbįb Mõųşşã ĄhmąţPas encore d'évaluation
- Groupe 8 Protéines MembranaireDocument4 pagesGroupe 8 Protéines MembranaireLounes MazariPas encore d'évaluation
- 2eme Cour PhysiologieDocument8 pages2eme Cour Physiologiederdiri imènePas encore d'évaluation
- Cours Membrane Plasmique & Hyaloplasme Microsoft Office WordDocument19 pagesCours Membrane Plasmique & Hyaloplasme Microsoft Office WordJoan Jean Jephthé Babo100% (4)
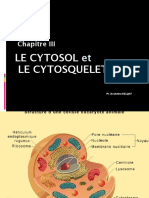
- CH3 - Cytosol-Cytosquelette - B.BDocument47 pagesCH3 - Cytosol-Cytosquelette - B.Bone asslanPas encore d'évaluation
- Membrane PlasmiqueDocument17 pagesMembrane PlasmiqueSami SamirPas encore d'évaluation
- La Membrane Plasmique: I. GénéralitésDocument17 pagesLa Membrane Plasmique: I. GénéralitésAisSa KeRrøùùmPas encore d'évaluation
- TD N °2 de Cytologie - Membrane Plasmique 2023-2024Document4 pagesTD N °2 de Cytologie - Membrane Plasmique 2023-2024Yassine YacinePas encore d'évaluation
- Cours 3. Membrane PlasmiqueDocument14 pagesCours 3. Membrane PlasmiqueNã BîlPas encore d'évaluation
- SUPPORT UE1B-KREJBICH-Homéostasie 2-17.10.22Document39 pagesSUPPORT UE1B-KREJBICH-Homéostasie 2-17.10.22Yann legendrePas encore d'évaluation
- Biologie Cellulaire CHAPITRE 3Document3 pagesBiologie Cellulaire CHAPITRE 3rida ElqodsPas encore d'évaluation
- Partie 2. Cours de BioCell 1ASNVDocument17 pagesPartie 2. Cours de BioCell 1ASNVايمن معطااللهPas encore d'évaluation
- Sujeujet Bio Celle Session 2 PCEM1Document4 pagesSujeujet Bio Celle Session 2 PCEM1medPas encore d'évaluation
- MPDFDocument4 pagesMPDFBad MaxPas encore d'évaluation
- Epreuve Bio Cell - Numerus - 2021-2022Document3 pagesEpreuve Bio Cell - Numerus - 2021-2022Mohamed DickoPas encore d'évaluation
- Structure Et Physiologie Des Eucaryotes 2018-2019-L2S4Document7 pagesStructure Et Physiologie Des Eucaryotes 2018-2019-L2S4Original ZeyePas encore d'évaluation
- La MembraneDocument23 pagesLa MembraneFanny GrouxPas encore d'évaluation
- Chapitre Vi Paroi PectocellulosiqueDocument9 pagesChapitre Vi Paroi Pectocellulosiqueleonce sedjroPas encore d'évaluation
- La Membrane PlasmiqueDocument53 pagesLa Membrane PlasmiqueahlemmPas encore d'évaluation
- Chapitre V La Membrane PlasmiqueDocument7 pagesChapitre V La Membrane PlasmiqueHatim Aitlaama100% (1)
- Electrophysiologie Membranaire 2020Document11 pagesElectrophysiologie Membranaire 2020Lina AmiraPas encore d'évaluation
- 4 - 1 Membrane PlasmiqueDocument43 pages4 - 1 Membrane PlasmiqueCelia Andreu AliagaPas encore d'évaluation
- Biologie CellulaireDocument37 pagesBiologie Cellulairenihalaouam05Pas encore d'évaluation
- CH I Indroduction À La Biotechnologie MOUELLEF A 2021Document12 pagesCH I Indroduction À La Biotechnologie MOUELLEF A 2021Idrissa SINONPas encore d'évaluation
- Filiere: Sciences de La Vie MODULE: Biologie Des Organismes Végétaux (BOV) Partie Thallophytes: Les ChampignonsDocument26 pagesFiliere: Sciences de La Vie MODULE: Biologie Des Organismes Végétaux (BOV) Partie Thallophytes: Les ChampignonsMohammed BenyachiPas encore d'évaluation
- UE2 La-Membrane-PlasmiqueDocument17 pagesUE2 La-Membrane-Plasmiquesun-nee-chan9Pas encore d'évaluation
- Jean Marc Governatori: Vers Une Santé TotaleDocument27 pagesJean Marc Governatori: Vers Une Santé TotaleAlliance Ecologiste IndépendantePas encore d'évaluation
- QCMDocument13 pagesQCMNassim OuabouPas encore d'évaluation
- ED Milieu Interieur CorrectionDocument5 pagesED Milieu Interieur CorrectionBouabidi100% (4)
- Biologie VegetaleDocument14 pagesBiologie Vegetaleblogmalika100% (1)
- Cours mtt1 l1 Snvdefinitif-2 PDFDocument30 pagesCours mtt1 l1 Snvdefinitif-2 PDFFarah TouatiPas encore d'évaluation
- Nutrition en CalciumDocument11 pagesNutrition en CalciumGladio SeifPas encore d'évaluation
- La Mitose MouradDocument5 pagesLa Mitose MouradMourad Ben Henda100% (2)
- 1ac AdilDocument2 pages1ac AdilAdil Adil0% (1)
- Secrets Éternels de Santé Et de RajeunissementDocument26 pagesSecrets Éternels de Santé Et de RajeunissementLadeu Kone100% (1)
- Présentation 8Document8 pagesPrésentation 8Youssra AzPas encore d'évaluation
- 2 - La Cellule Végétale - FicheDocument16 pages2 - La Cellule Végétale - FicheLisa FoucherPas encore d'évaluation
- Annale - BiologieDocument71 pagesAnnale - BiologieDesakoBEATSPas encore d'évaluation
- Chap II Récepteurs Membranaires Et Molécules de Signalisation IntracellulaireDocument28 pagesChap II Récepteurs Membranaires Et Molécules de Signalisation IntracellulaireAbdoulaye sebyPas encore d'évaluation
- Troubles Du Metabolisme Des Lipides: DR Chabane SariDocument30 pagesTroubles Du Metabolisme Des Lipides: DR Chabane Sarimohamed rayanPas encore d'évaluation
- 04-BP1 BP2-Coiffure-Biologie AppliquéeDocument16 pages04-BP1 BP2-Coiffure-Biologie AppliquéeThierry Pano Doubleface DucosPas encore d'évaluation
- Cours TIA - Chapitre 2 PDFDocument11 pagesCours TIA - Chapitre 2 PDFHoussine GuePas encore d'évaluation
- Lexique GenetiqueDocument4 pagesLexique GenetiqueNeginow 627Pas encore d'évaluation
- Raw Milk FloraDocument134 pagesRaw Milk FloraDCPas encore d'évaluation
- TP5 Tissu SanguinDocument8 pagesTP5 Tissu SanguinNihal KrikaPas encore d'évaluation
- Biologie MoleculaireDocument20 pagesBiologie MoleculaireKhalid ChrisPas encore d'évaluation
- SCH Cours Partie 1 (Parasites) FRDocument46 pagesSCH Cours Partie 1 (Parasites) FRIrnatene ChahinezPas encore d'évaluation
- Unité D'enseignement L3 (S5+S6)Document5 pagesUnité D'enseignement L3 (S5+S6)Imene BaghaliPas encore d'évaluation
- Sujet Biologie FMSP 2010Document2 pagesSujet Biologie FMSP 2010Bile DjetouanPas encore d'évaluation
- ?physiologie ?Document67 pages?physiologie ?Espoir EDOHPas encore d'évaluation
- Lésions Élémentaires CellulaireDocument77 pagesLésions Élémentaires CellulaireLaMri IkRamPas encore d'évaluation
- Organismes Uni Et Multi PDFDocument27 pagesOrganismes Uni Et Multi PDFFahroudinePas encore d'évaluation
- DS 6 522 2022-2023 26Document3 pagesDS 6 522 2022-2023 26Odunayo Onésime100% (1)