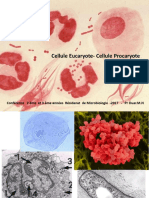
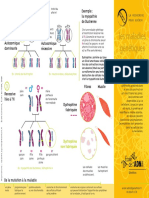
Vous aimerez peut-être aussi
- Membranes cellulaires: Les Grands Articles d'UniversalisD'EverandMembranes cellulaires: Les Grands Articles d'UniversalisPas encore d'évaluation
- BIO 111 - CHAP - VI - VII - Et - VIIIDocument9 pagesBIO 111 - CHAP - VI - VII - Et - VIIIBerthe Aristide Ngue NkongoPas encore d'évaluation
- Champignons: Les Grands Articles d'UniversalisD'EverandChampignons: Les Grands Articles d'UniversalisPas encore d'évaluation
- CytologieDocument43 pagesCytologieMouats Badreddine100% (2)
- Chapitre I Structure de La CelluleDocument107 pagesChapitre I Structure de La CelluleDescartes KpannePas encore d'évaluation
- TD7 Les Organites de La Cellule PDFDocument42 pagesTD7 Les Organites de La Cellule PDFZineb DahmaniPas encore d'évaluation
- Anatomie 1 (Récupération Automatique)Document3 pagesAnatomie 1 (Récupération Automatique)Hossam AnaPas encore d'évaluation
- Cours Membrane Plasmique & Hyaloplasme Microsoft Office WordDocument19 pagesCours Membrane Plasmique & Hyaloplasme Microsoft Office WordJoan Jean Jephthé Babo100% (4)
- Cours Biologie Stu s1 StuDocument27 pagesCours Biologie Stu s1 StuDef CamPas encore d'évaluation
- Anatomie Avant Dernière PartieDocument9 pagesAnatomie Avant Dernière PartieAdn AdnPas encore d'évaluation
- Cours de Microbiologie 2eme Année BiologieDocument13 pagesCours de Microbiologie 2eme Année BiologieKàdî David100% (11)
- La CelluleDocument26 pagesLa CellulesaadiaPas encore d'évaluation
- 1 Partie-1 Cours-S1 Introduction Const-ChimiquesDocument50 pages1 Partie-1 Cours-S1 Introduction Const-Chimiquesعالم البحث Monde de la recherchePas encore d'évaluation
- Cours de Bactériologie Chap 1 Description de La Cellule BactérienneDocument29 pagesCours de Bactériologie Chap 1 Description de La Cellule BactérienneSOKAMTE TEGANG AlphonsePas encore d'évaluation
- La Cellule PDFDocument4 pagesLa Cellule PDFnehemie nwayPas encore d'évaluation
- Structure Des Bactéries 2018 2019Document8 pagesStructure Des Bactéries 2018 2019Ismail HamrouniPas encore d'évaluation
- 01 - A-Structure Et Architecture de La MembraneDocument41 pages01 - A-Structure Et Architecture de La MembraneKhelifi SarraPas encore d'évaluation
- l12 Microbio Chap1Document9 pagesl12 Microbio Chap1Alhassane BahPas encore d'évaluation
- 2-Cellule Ses ConstituantsDocument56 pages2-Cellule Ses ConstituantsAy ManPas encore d'évaluation
- Chapitre 1. Cellule Procaryote 1Document4 pagesChapitre 1. Cellule Procaryote 1lynda divine EmièmePas encore d'évaluation
- La CelluleDocument6 pagesLa Celluleikram dahdouhPas encore d'évaluation
- 3.schema General Des Cellules Procaryotes Et Eucaryotes 2021-2022Document56 pages3.schema General Des Cellules Procaryotes Et Eucaryotes 2021-2022Le BoyaïstePas encore d'évaluation
- La CelluleDocument52 pagesLa Cellulesimo.elhaouate.1000Pas encore d'évaluation
- 01 Generalites Sur La CelluleDocument11 pages01 Generalites Sur La CelluleAbdoul karim SamakePas encore d'évaluation
- 2021 - Cours La Cellule Version ComplèteDocument10 pages2021 - Cours La Cellule Version Complèteagathe surusPas encore d'évaluation
- Cour 1 CytoDocument4 pagesCour 1 CytoSerine LaraPas encore d'évaluation
- Membrane PlasmiqueDocument7 pagesMembrane PlasmiqueSara BoudjemaiPas encore d'évaluation
- Chapitre II La Cellule BactérienneDocument22 pagesChapitre II La Cellule BactérienneMouhcin BelmekiPas encore d'évaluation
- Capture D'écran . 2024-01-07 À 01.16.35Document23 pagesCapture D'écran . 2024-01-07 À 01.16.35inesbilliePas encore d'évaluation
- La CelluleDocument6 pagesLa CelluleHossam AnaPas encore d'évaluation
- Structure BactérienneDocument72 pagesStructure BactériennekhouildiPas encore d'évaluation
- Chap 2 MPDocument7 pagesChap 2 MPdramohandlhadjPas encore d'évaluation
- Chapitre1 LacellulecoursDocument6 pagesChapitre1 LacellulecoursHéloïse BlrPas encore d'évaluation
- BacterieDocument7 pagesBacterieSalah KhaldiPas encore d'évaluation
- Chapitre II: Morphologie Et Structure de La Cellule Bacterienne I. Technique D'étudeDocument8 pagesChapitre II: Morphologie Et Structure de La Cellule Bacterienne I. Technique D'étudeWided HamdiPas encore d'évaluation
- 02 Revetement-CellulaireDocument6 pages02 Revetement-CellulaireSagno AgnèsPas encore d'évaluation
- 9 CLASSE DE Tle C-1Document103 pages9 CLASSE DE Tle C-1Njiasse MaazouPas encore d'évaluation
- Chapitre VIDocument6 pagesChapitre VISiremPas encore d'évaluation
- Chap 01Document33 pagesChap 01hamidaPas encore d'évaluation
- Biologie C - Séance 2Document56 pagesBiologie C - Séance 2AymanePas encore d'évaluation
- Chapitre 2 Anatomie BacterienneDocument21 pagesChapitre 2 Anatomie BacteriennefsxnsstoaehkbhlzygPas encore d'évaluation
- Bio CellDocument23 pagesBio Cellkouki samarPas encore d'évaluation
- Biocellsession 2Document15 pagesBiocellsession 2Eroline GuelciPas encore d'évaluation
- Structure Et Anatomie Fonctionnelle Des Bacteries-ConvertiDocument55 pagesStructure Et Anatomie Fonctionnelle Des Bacteries-ConvertiAmiraBenhammou100% (1)
- Structure Et Anatomie Fonctionnelle Des BacteriesDocument55 pagesStructure Et Anatomie Fonctionnelle Des BacteriesAmira BenhammouPas encore d'évaluation
- Cellule, Structure Et FonctionDocument7 pagesCellule, Structure Et Fonctionsokhnatouba99Pas encore d'évaluation
- Structure Des BactériesDocument9 pagesStructure Des BactérieskhouildiPas encore d'évaluation
- TD Biologie Cellulaire 2024Document4 pagesTD Biologie Cellulaire 2024Halima MintPas encore d'évaluation
- Microbiologie Générale Chap. II 2021Document12 pagesMicrobiologie Générale Chap. II 2021Sid AliPas encore d'évaluation
- Cours Biologie Cellulaire MELDocument5 pagesCours Biologie Cellulaire MELhanaPas encore d'évaluation
- Chapitre II Partie 2Document14 pagesChapitre II Partie 2KahinaPas encore d'évaluation
- Chapitre 1Document5 pagesChapitre 1Fatïma ZohraPas encore d'évaluation
- La Cellule EucaryoteDocument2 pagesLa Cellule EucaryoteIngrid Marchaux100% (1)
- Chapitre 2 Anatomie BacterienneDocument20 pagesChapitre 2 Anatomie BacterienneLamia HamdiPas encore d'évaluation
- Identite BiologiaueDocument15 pagesIdentite BiologiauePessu EricPas encore d'évaluation
- Chap - 6 - Les Compartiments IntracellulairesDocument4 pagesChap - 6 - Les Compartiments IntracellulairesMoeanau GrangerPas encore d'évaluation
- Ribosome 2021 - DR Zergui ADocument17 pagesRibosome 2021 - DR Zergui Amalak zerizerPas encore d'évaluation
- Introduction A La Bio CellDocument15 pagesIntroduction A La Bio CellLilou GrambertPas encore d'évaluation
- Sujeujet Bio Celle Session 2 PCEM1Document4 pagesSujeujet Bio Celle Session 2 PCEM1medPas encore d'évaluation
- Chapitre II BactériesDocument30 pagesChapitre II Bactériesislam.brm1332Pas encore d'évaluation
- Cours1.sang GeneralitesDocument120 pagesCours1.sang GeneralitesTemtem HmPas encore d'évaluation
- TD Solution...Document15 pagesTD Solution...Mohamede KhamlichiPas encore d'évaluation
- 1 Chap 1 Monde Microbien PRDocument79 pages1 Chap 1 Monde Microbien PRAyatePas encore d'évaluation
- Génétique ExamensDocument30 pagesGénétique ExamensTemtem HmPas encore d'évaluation
- Exercices Corrig S - G N Tique HumaineDocument7 pagesExercices Corrig S - G N Tique HumaineBelgasem Assel100% (1)
- Le Gros Arbre Qui Cachait La ForêtDocument3 pagesLe Gros Arbre Qui Cachait La ForêtTemtem HmPas encore d'évaluation
- Régulation de La Masse Osseuse Par La LeptineDocument6 pagesRégulation de La Masse Osseuse Par La LeptineTemtem HmPas encore d'évaluation
- Contrôle Neural de Remodelage Osseux Le Rôle Crucial de La LeptineDocument2 pagesContrôle Neural de Remodelage Osseux Le Rôle Crucial de La LeptineTemtem HmPas encore d'évaluation
- Ecologie Generale Ii - Td1 Exercice IDocument2 pagesEcologie Generale Ii - Td1 Exercice IAicha GHOUZLANIPas encore d'évaluation
- Contrôle Neural de Remodelage Osseux Le Rôle Crucial de La LeptineDocument2 pagesContrôle Neural de Remodelage Osseux Le Rôle Crucial de La LeptineTemtem HmPas encore d'évaluation
- Thèse Leptine 2Document142 pagesThèse Leptine 2Temtem HmPas encore d'évaluation
- Cours de Biochimie: Enzymologie ApprofondieDocument90 pagesCours de Biochimie: Enzymologie ApprofondieTemtem HmPas encore d'évaluation
- Régulation de La Masse Osseuse Par La LeptineDocument6 pagesRégulation de La Masse Osseuse Par La LeptineTemtem HmPas encore d'évaluation
- Le Gros Arbre Qui Cachait La ForêtDocument3 pagesLe Gros Arbre Qui Cachait La ForêtTemtem HmPas encore d'évaluation
- SalmonellaDocument47 pagesSalmonellaTarik DamirPas encore d'évaluation
- Prise en Charge de La Gangrene de Fournier: Experience de L Hopital de SikassoDocument8 pagesPrise en Charge de La Gangrene de Fournier: Experience de L Hopital de SikassoIJAR JOURNALPas encore d'évaluation
- Physiopathologie Des OedémesDocument23 pagesPhysiopathologie Des OedémesBra BenzPas encore d'évaluation
- BERTHELEMYDocument22 pagesBERTHELEMYibrahima diengPas encore d'évaluation
- Pharmacologie Des Anti UlcéreuxDocument5 pagesPharmacologie Des Anti UlcéreuxNada KarachiPas encore d'évaluation
- RapportdestageDocument14 pagesRapportdestageCheikh Ismaïla BAPas encore d'évaluation
- Blandine CsDocument2 pagesBlandine CsSALIHOUPas encore d'évaluation
- Les VibrationsDocument2 pagesLes VibrationsDi BenamoPas encore d'évaluation
- A Propos de La Névrose Obsessionnelle FéminineDocument12 pagesA Propos de La Névrose Obsessionnelle FéminineFrixos MaraveliasPas encore d'évaluation
- DSH AIGUE Extrait Du Livre Urgences PediatriquesDocument9 pagesDSH AIGUE Extrait Du Livre Urgences Pediatriquesptit formatPas encore d'évaluation
- Carcinome Papillaire Intra-Kystique Du Sein: Quelle Indication de La Radiotherapie? A Travers Un Cas Clinique Et Une Revue de La LitteratureDocument4 pagesCarcinome Papillaire Intra-Kystique Du Sein: Quelle Indication de La Radiotherapie? A Travers Un Cas Clinique Et Une Revue de La LitteratureIJAR JOURNALPas encore d'évaluation
- Myélome - PrésentationDocument23 pagesMyélome - PrésentationLouisGrassoPas encore d'évaluation
- 11 Maladies GenetiquesDocument2 pages11 Maladies GenetiquesThéodore KONDIPas encore d'évaluation
- Diapo 1 - Notions GénéralesDocument24 pagesDiapo 1 - Notions GénéralesAdila DjelailiaPas encore d'évaluation
- Cacao CultureDocument24 pagesCacao CultureJephte KenfackPas encore d'évaluation
- Sémiologie de L'appareil Respiratoire PDFDocument16 pagesSémiologie de L'appareil Respiratoire PDFanon_989228910Pas encore d'évaluation
- 3) Malformations de La Charnière Occipito-VertébraleDocument5 pages3) Malformations de La Charnière Occipito-VertébralehoudaPas encore d'évaluation
- Lautorite Du ChretienDocument5 pagesLautorite Du Chretienkoffi serge pacome MBRAPas encore d'évaluation
- Thérapeutiques Dentino-PulpairesDocument38 pagesThérapeutiques Dentino-PulpairesKaoutar KbPas encore d'évaluation
- Curiosités de La GénétiqueDocument4 pagesCuriosités de La GénétiqueAlejandroMarínPas encore d'évaluation
- 20 La Sero Prevalence de L'hepatiteDocument75 pages20 La Sero Prevalence de L'hepatiteAthar SoudaniPas encore d'évaluation
- These: Universite de MontpellierDocument203 pagesThese: Universite de Montpelliertaoufik akabliPas encore d'évaluation
- Catalogue Cursri Ifpek RennesDocument83 pagesCatalogue Cursri Ifpek RennesloredanatudorachePas encore d'évaluation
- 3.résumé Hérédité HumaineDocument3 pages3.résumé Hérédité HumaineAyoub LilouPas encore d'évaluation
- Position Officielle de L'association Américaine de Diététique Et Des Diététiciens Canadiens Au Sujet de L'alimentation VégétarienneDocument18 pagesPosition Officielle de L'association Américaine de Diététique Et Des Diététiciens Canadiens Au Sujet de L'alimentation Végétarienneveronique henryPas encore d'évaluation
- Memoire Fin D'etude Ratard de RéveilDocument67 pagesMemoire Fin D'etude Ratard de RéveilBessmaPas encore d'évaluation
- Étude Comparéechine-FranceDocument26 pagesÉtude Comparéechine-FranceDidier CoutanceauPas encore d'évaluation
- MEP D'anatomie III (CM + TP)Document65 pagesMEP D'anatomie III (CM + TP)Anass VillaPas encore d'évaluation
- Chaines Legeres LibresDocument3 pagesChaines Legeres LibresNabil BahouraPas encore d'évaluation
- ChronDocument8 pagesChronViorel BudacPas encore d'évaluation
- Manuel pour les débutants Fabriquez des savons naturelsD'EverandManuel pour les débutants Fabriquez des savons naturelsÉvaluation : 3 sur 5 étoiles3/5 (2)
- Cahier de jeux de stimulation cognitive: Sujets Alzheimer, désorientés, démences, amnésiesD'EverandCahier de jeux de stimulation cognitive: Sujets Alzheimer, désorientés, démences, amnésiesPas encore d'évaluation
- La thérapie libre: La Thérapie de LIBération et Résilience EmotionnelleD'EverandLa thérapie libre: La Thérapie de LIBération et Résilience EmotionnelleÉvaluation : 5 sur 5 étoiles5/5 (1)
- Géologie de l'Amérique: Les Grands Articles d'UniversalisD'EverandGéologie de l'Amérique: Les Grands Articles d'UniversalisPas encore d'évaluation
- La vie des abeilles: Prix Nobel de littératureD'EverandLa vie des abeilles: Prix Nobel de littératureÉvaluation : 4 sur 5 étoiles4/5 (41)
- Améliorer votre mémoire: Un Guide pour l'augmentation de la puissance du cerveau, utilisant des techniques et méthodesD'EverandAméliorer votre mémoire: Un Guide pour l'augmentation de la puissance du cerveau, utilisant des techniques et méthodesÉvaluation : 5 sur 5 étoiles5/5 (2)
- Harmonisation Energétique des Lieux: Habitat et haut-lieux sacrés 2020D'EverandHarmonisation Energétique des Lieux: Habitat et haut-lieux sacrés 2020Évaluation : 2.5 sur 5 étoiles2.5/5 (3)
- Histoire de la psychologie scientifique: De la naissance de la psychologie à la neuropsychologie et aux champs d'application les plus actuelsD'EverandHistoire de la psychologie scientifique: De la naissance de la psychologie à la neuropsychologie et aux champs d'application les plus actuelsPas encore d'évaluation
- Les 10 Secrets pour une Vie Plus Heureuse avec la Maladie de ParkinsonD'EverandLes 10 Secrets pour une Vie Plus Heureuse avec la Maladie de ParkinsonPas encore d'évaluation
- Microbiologie médicale I: agents pathogènes et microbiome humainD'EverandMicrobiologie médicale I: agents pathogènes et microbiome humainÉvaluation : 5 sur 5 étoiles5/5 (1)
- Le CODE DE DIEU: Le secret de notre passé, la promesse de notre avenirD'EverandLe CODE DE DIEU: Le secret de notre passé, la promesse de notre avenirÉvaluation : 4 sur 5 étoiles4/5 (23)
- Qu'arrive t'il a la terre arable ?: Le problème de l'érosion du sol.D'EverandQu'arrive t'il a la terre arable ?: Le problème de l'érosion du sol.Évaluation : 5 sur 5 étoiles5/5 (1)
- Atlas permanent de l'Union européenne: 5e édition revue et augmentéeD'EverandAtlas permanent de l'Union européenne: 5e édition revue et augmentéePas encore d'évaluation
- La Conscience Et L'Univers Existent Sans Commencement Ni FinD'EverandLa Conscience Et L'Univers Existent Sans Commencement Ni FinPas encore d'évaluation
- Les Graines d'Abricot - Remède de Cancer avec de la Vitamine B17 ?: La Médecine Antique Que l'Industrie Pharmaceutique Moderne CacheD'EverandLes Graines d'Abricot - Remède de Cancer avec de la Vitamine B17 ?: La Médecine Antique Que l'Industrie Pharmaceutique Moderne CachePas encore d'évaluation
- Pourquoi suis-je différent?: Accepter et découvrir la puissance de l'hypersensibilitéD'EverandPourquoi suis-je différent?: Accepter et découvrir la puissance de l'hypersensibilitéPas encore d'évaluation
- Transition énergétique ET Changement climatique: Développements et Perspectives d'AvenirD'EverandTransition énergétique ET Changement climatique: Développements et Perspectives d'AvenirPas encore d'évaluation
- Un vent de folie: L'éolien en France : mensonge et arnaque ?D'EverandUn vent de folie: L'éolien en France : mensonge et arnaque ?Pas encore d'évaluation
- Connaître ses réactions émotionnelles, pour maîtriser ses émotionsD'EverandConnaître ses réactions émotionnelles, pour maîtriser ses émotionsPas encore d'évaluation
- Traitement des psychonévroses: par la rééducation du contrôle cérébralD'EverandTraitement des psychonévroses: par la rééducation du contrôle cérébralPas encore d'évaluation