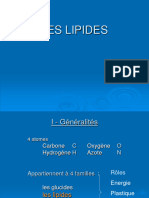
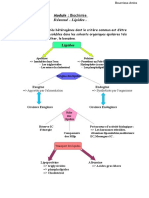
Vous aimerez peut-être aussi
- Les LipidesDocument8 pagesLes Lipidesyousraghanem23Pas encore d'évaluation
- Cours Lipides 20 21Document16 pagesCours Lipides 20 21oumayma salihiPas encore d'évaluation
- 1 - Les LipidesDocument25 pages1 - Les LipidesFanny GrouxPas encore d'évaluation
- ACIDE GRAS - EtudiantsDocument48 pagesACIDE GRAS - EtudiantsZaki ByzPas encore d'évaluation
- Cours Lipides 1 2023Document22 pagesCours Lipides 1 2023Amani Mann'fangan EphremePas encore d'évaluation
- LIPIDES Biochimie StructuraleDocument82 pagesLIPIDES Biochimie StructuraleCheick SANOUPas encore d'évaluation
- Structure Des Lipides Medecine DentaireDocument93 pagesStructure Des Lipides Medecine DentairezadiPas encore d'évaluation
- LIPIDES Biochimie Structurale-1Document69 pagesLIPIDES Biochimie Structurale-1Cheick SANOUPas encore d'évaluation
- LipideDocument28 pagesLipideChanez FekirPas encore d'évaluation
- Resume Les LipidesDocument14 pagesResume Les LipidesihcPas encore d'évaluation
- Lipids (Part 1)Document33 pagesLipids (Part 1)Deakanohv JeanPas encore d'évaluation
- 1 1 Structure Des Lipides PDFDocument12 pages1 1 Structure Des Lipides PDFNaouel BoudPas encore d'évaluation
- Compostion +proprietes Des Corps Gras-5 by Nacerinho Dhiya EddineDocument4 pagesCompostion +proprietes Des Corps Gras-5 by Nacerinho Dhiya EddineMadjidAvengersPas encore d'évaluation
- Chapitre 1-3 Les LipidesDocument17 pagesChapitre 1-3 Les Lipidesmohamadou moustapha gayePas encore d'évaluation
- Aliments Simples Cours2 Theme1Document6 pagesAliments Simples Cours2 Theme1Amor FaouziPas encore d'évaluation
- 2 - LipidesDocument35 pages2 - LipidesRima AllouniPas encore d'évaluation
- 2-Résumé Bournisa Amira 2015-2016Document25 pages2-Résumé Bournisa Amira 2015-2016Farouk DjenienPas encore d'évaluation
- Lipides de Stockage Lipides de Structure Vitamines Liposolubles Pigments Hormones Agent Émulsifiant Médiateur de L'inflammationDocument19 pagesLipides de Stockage Lipides de Structure Vitamines Liposolubles Pigments Hormones Agent Émulsifiant Médiateur de L'inflammationJustin DrouetPas encore d'évaluation
- Présentation 3Document32 pagesPrésentation 3Yahia BerbouchiPas encore d'évaluation
- Metabolisme Des LipidesDocument150 pagesMetabolisme Des LipidesKdimosaure KdimoPas encore d'évaluation
- Lecon 2 - Les Macomolecules Biologiques - Dec13Document43 pagesLecon 2 - Les Macomolecules Biologiques - Dec13yannicknzigangPas encore d'évaluation
- Glucides 1Document47 pagesGlucides 1Rania MaddahPas encore d'évaluation
- Cours - Biochimie Structurale - Lipides - CAG - UNA - 2019 - CopieDocument83 pagesCours - Biochimie Structurale - Lipides - CAG - UNA - 2019 - CopieStellaPas encore d'évaluation
- Cours Lipides Structurale (Compatibility Mode)Document80 pagesCours Lipides Structurale (Compatibility Mode)bikungracePas encore d'évaluation
- Chapitre II Les LipidesDocument7 pagesChapitre II Les LipidesEssassi AmmarPas encore d'évaluation
- Seance 5Document17 pagesSeance 5Pascal GadedjissoPas encore d'évaluation
- 08.5-Les Lipides SimplesDocument8 pages08.5-Les Lipides SimplesBook HunterPas encore d'évaluation
- 2-Lipides ComplexesDocument48 pages2-Lipides Complexes130660LYC OMAR BENABDELAZIZ NEDROMAPas encore d'évaluation
- 2-Lipides ComplexesDocument48 pages2-Lipides Complexes130660LYC OMAR BENABDELAZIZ NEDROMAPas encore d'évaluation
- Biochimie FDocument7 pagesBiochimie FMathin BrouPas encore d'évaluation
- Les Corps Gras AlimentairesDocument18 pagesLes Corps Gras AlimentairesZineb BelalaPas encore d'évaluation
- Matière Vivante-1Document53 pagesMatière Vivante-1malomedPas encore d'évaluation
- Cours de Biochimie Structurale 2022 2023 LIPIDEDocument25 pagesCours de Biochimie Structurale 2022 2023 LIPIDEvivaspro900Pas encore d'évaluation
- COURS LIPIDES FMS 2022 - 2023 (Pr. SAID)Document19 pagesCOURS LIPIDES FMS 2022 - 2023 (Pr. SAID)Abdelhamid ArouiPas encore d'évaluation
- Lipides Cours 01 PDFDocument26 pagesLipides Cours 01 PDFpublicité en flopOfficielPas encore d'évaluation
- Metab Des Lipides Boukhira 2018 19-1Document145 pagesMetab Des Lipides Boukhira 2018 19-1Cheick KonePas encore d'évaluation
- Lip Final NewDocument70 pagesLip Final NewAymanePas encore d'évaluation
- Biochimie Structurale ESTBADocument79 pagesBiochimie Structurale ESTBAtolisso.yawoviPas encore d'évaluation
- 2-Polycopie LipidesDocument27 pages2-Polycopie Lipidesyanis100% (1)
- LipidesDocument42 pagesLipidesKEBIR ZAHRAPas encore d'évaluation
- Les Corps Gras AlimentairesDocument14 pagesLes Corps Gras AlimentairesmsitankoPas encore d'évaluation
- Fiches de Révisions Biologie Psycho L1Document16 pagesFiches de Révisions Biologie Psycho L1MilaPas encore d'évaluation
- Lip IdesDocument8 pagesLip IdesYousra INDIAPas encore d'évaluation
- Lipides 2Document12 pagesLipides 2Hadrien De GreefPas encore d'évaluation
- Notions Fondamentales Sur Corps GrasDocument14 pagesNotions Fondamentales Sur Corps GrasRayène BekirPas encore d'évaluation
- Copie de Lipides Diapos 1Document59 pagesCopie de Lipides Diapos 1Maha AzzouziPas encore d'évaluation
- pb9 Molécules d' Intérêt Biologique, DocumentsDocument5 pagespb9 Molécules d' Intérêt Biologique, Documentsantony.assantePas encore d'évaluation
- Hininger Favier Isabelle P01Document72 pagesHininger Favier Isabelle P01ADRIANPas encore d'évaluation
- Lipides 2Document23 pagesLipides 2ghita.apu100% (1)
- Cours Complet D'svteehb Apc-1Document61 pagesCours Complet D'svteehb Apc-1abdoulbenrazadPas encore d'évaluation
- 3 Lipides CoursDocument7 pages3 Lipides CoursZaki ByzPas encore d'évaluation
- Introduction Sur Le Métabolisme LipidesDocument21 pagesIntroduction Sur Le Métabolisme Lipidescep100% (1)
- Cours de Biologie Cellulaire - CHP IDocument18 pagesCours de Biologie Cellulaire - CHP ISwayli HossamPas encore d'évaluation
- 07 LipidesDocument81 pages07 LipidesSaida AdjemiPas encore d'évaluation
- Chapitre I Composition Chimique de La Cellule 1 (2) - Copie NEDocument46 pagesChapitre I Composition Chimique de La Cellule 1 (2) - Copie NEkhalid BOUBKRIPas encore d'évaluation
- Composition Elementaire de La Matiere Vivante PDFDocument18 pagesComposition Elementaire de La Matiere Vivante PDFpbombartPas encore d'évaluation
- Les LipidesDocument17 pagesLes Lipidesneyrine atmPas encore d'évaluation
- Biologie Preparation Concours ZoéDocument32 pagesBiologie Preparation Concours ZoéJoe Zoe Francescodash MukokoPas encore d'évaluation
- Polycope Lipides 1ére Année Médecine PDFDocument31 pagesPolycope Lipides 1ére Année Médecine PDFKacem Kamli100% (1)
- 70 Examens Corriges Chimie Organique Generale SMC s3 2014Document12 pages70 Examens Corriges Chimie Organique Generale SMC s3 2014r.agadimPas encore d'évaluation
- Etude BibliographiqueDocument23 pagesEtude Bibliographiquemliki mohamed100% (1)
- Mémoire Version FinaleDocument61 pagesMémoire Version FinaleSafae hirak100% (1)
- Classification LCPC Du Sol MdsDocument5 pagesClassification LCPC Du Sol MdsDj'aaFer Wafo100% (3)
- Compte Rendu TP Physico Chimie Des PolyméresDocument18 pagesCompte Rendu TP Physico Chimie Des PolyméresNozha HammamiPas encore d'évaluation
- Tensioactivité 1 Chimie Surfaces 3A FondamentaleDocument6 pagesTensioactivité 1 Chimie Surfaces 3A FondamentaleZahra l زهرةPas encore d'évaluation
- Chapitre 1Document6 pagesChapitre 1riham rihamPas encore d'évaluation
- Chapitre 8Document41 pagesChapitre 8Gabriel Lapointe-GuayPas encore d'évaluation
- 1 Terminale D ChimieDocument3 pages1 Terminale D ChimieJoel AsuraPas encore d'évaluation
- 1cm3uifap 891539Document4 pages1cm3uifap 891539GueyePas encore d'évaluation
- Shampooings - Composition Et Caractéristiques de La Base LavanteDocument15 pagesShampooings - Composition Et Caractéristiques de La Base LavanteMohamed KARASAD100% (1)
- Pétro-Chimie MorouDocument6 pagesPétro-Chimie Morouharouna souley hegaPas encore d'évaluation
- Micro Biolog I eDocument4 pagesMicro Biolog I eChescoPas encore d'évaluation
- Chimie 2Document3 pagesChimie 2Elvis Wilfried PossiPas encore d'évaluation
- Structure Et Métabolisme HémoglobinE - VFDocument46 pagesStructure Et Métabolisme HémoglobinE - VFs22g9ndkwjPas encore d'évaluation
- 2021 Centres Etrangers ExoB Sujet EsterRhum 5ptsDocument3 pages2021 Centres Etrangers ExoB Sujet EsterRhum 5ptsRaphael DEMIANPas encore d'évaluation
- Ahlam LoucifDocument64 pagesAhlam Louciflalia djekidelPas encore d'évaluation
- Les HalogènesDocument35 pagesLes HalogènesKrim Issam Eddine100% (4)
- Mémoire: Evaluation de L'activité Antioxydante Des Extraits deDocument52 pagesMémoire: Evaluation de L'activité Antioxydante Des Extraits deChouaibi FathiaPas encore d'évaluation
- BAC 2022 Spécialité SVT Polynésie - Jour 2Document7 pagesBAC 2022 Spécialité SVT Polynésie - Jour 2LETUDIANT100% (1)
- Catalogue MCI 2019Document24 pagesCatalogue MCI 2019Nirina AndriaPas encore d'évaluation
- Contrôle de Biochimie Structurale WordDocument7 pagesContrôle de Biochimie Structurale WordNoorPas encore d'évaluation
- COURS Chim224 VG 2022-2023Document87 pagesCOURS Chim224 VG 2022-2023Salim SaoudPas encore d'évaluation
- Plantes (Dét)Document121 pagesPlantes (Dét)Laboratoire EL ALLALI - AGADIRPas encore d'évaluation
- Lipides Cours 01 PDFDocument26 pagesLipides Cours 01 PDFpublicité en flopOfficielPas encore d'évaluation
- Développement Des Leçons de La Série: Leçon 1 Le Vivant Et Le Non-VivantDocument38 pagesDéveloppement Des Leçons de La Série: Leçon 1 Le Vivant Et Le Non-VivantMoh'dTfailiPas encore d'évaluation
- 04 - Coumarines Et LignanesDocument14 pages04 - Coumarines Et LignanesAmira BenhammouPas encore d'évaluation
- Acides AminésDocument14 pagesAcides AminésA100% (1)
- Caractéristiques Des Eaux UséesDocument94 pagesCaractéristiques Des Eaux UséesMinouch K100% (1)
- TD Extraction Des Substances NaturellesDocument3 pagesTD Extraction Des Substances Naturellesnassima chenikhaPas encore d'évaluation