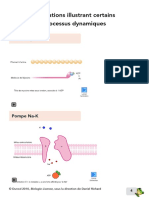
Vous aimerez peut-être aussi
- Les Enzymes de RestrictionDocument16 pagesLes Enzymes de RestrictionMadjidAvengersPas encore d'évaluation
- Chapitre11 Genie GenetiqueDocument8 pagesChapitre11 Genie GenetiqueSaid Sai100% (1)
- La Reparation de l'ADNDocument4 pagesLa Reparation de l'ADNŇøu ŞsāPas encore d'évaluation
- Cours Clonage Et Banque de GènesDocument9 pagesCours Clonage Et Banque de GènesMadjidAvengersPas encore d'évaluation
- Chapitre 3 Recombinaison GénétiqueDocument5 pagesChapitre 3 Recombinaison GénétiqueAbboud DjamaaPas encore d'évaluation
- Rappels Indispensables de GénétiqueDocument10 pagesRappels Indispensables de Génétiquebakiclio5Pas encore d'évaluation
- Medsci20153111p1014 5Document1 pageMedsci20153111p1014 5jaliyo8623Pas encore d'évaluation
- Microbiologie Partie 1Document31 pagesMicrobiologie Partie 1Hassan BouhoiiliPas encore d'évaluation
- SVT Devir n1 Cned 1Document4 pagesSVT Devir n1 Cned 1k. kiritoPas encore d'évaluation
- B-Constance Et Variation Du Dna - Mobilite PDFDocument17 pagesB-Constance Et Variation Du Dna - Mobilite PDFrhirhaPas encore d'évaluation
- Examen 28 Mai 2010-CorrigéDocument6 pagesExamen 28 Mai 2010-Corrigéikram bt0% (1)
- Chapitre IV VecteursDocument19 pagesChapitre IV Vecteursihcene aouesPas encore d'évaluation
- Definition UsthbDocument12 pagesDefinition UsthbMahieddine BoucekkinePas encore d'évaluation
- Questionnaire Bio 2023Document46 pagesQuestionnaire Bio 2023mpogazi clery arecPas encore d'évaluation
- Biochimie 08 - Réplication Et Réparation Des Erreurs AssociéesDocument14 pagesBiochimie 08 - Réplication Et Réparation Des Erreurs AssociéesNathan CohenPas encore d'évaluation
- 4 Genetique BacterienneDocument11 pages4 Genetique BacteriennemerzougPas encore d'évaluation
- 4 - Les VecteursDocument5 pages4 - Les VecteursChibname ChibnamePas encore d'évaluation
- UE1 Biologie Moleculaire Variation de L ADN 2010-2011Document59 pagesUE1 Biologie Moleculaire Variation de L ADN 2010-2011lobnaPas encore d'évaluation
- Chapitre 4Document27 pagesChapitre 4sabrinePas encore d'évaluation
- Chap - 2 - La Réplication de l'ADNDocument5 pagesChap - 2 - La Réplication de l'ADNDAHOUNOM Aboudou AzizouPas encore d'évaluation
- 5-Les Technologies de l'ADN RecombinantDocument7 pages5-Les Technologies de l'ADN RecombinantWi SsalPas encore d'évaluation
- Clonage PDFDocument37 pagesClonage PDFJalelo AbdelPas encore d'évaluation
- 15.02.2016 La RéplicationDocument10 pages15.02.2016 La RéplicationEaPas encore d'évaluation
- Polycopie Karouche 2Document163 pagesPolycopie Karouche 2Soraya YOUYOUPas encore d'évaluation
- Biologie Moléculaire Chapitre 1 T Procaryote M1Document8 pagesBiologie Moléculaire Chapitre 1 T Procaryote M1اميرة في عرش ابيPas encore d'évaluation
- Duplication de l'ADN - 011009Document65 pagesDuplication de l'ADN - 011009nonamethe427Pas encore d'évaluation
- Cours TD Gen ProDocument51 pagesCours TD Gen Prolobna67% (3)
- LA RÉPLICATION DE L'ADN - SVT 1èreDocument3 pagesLA RÉPLICATION DE L'ADN - SVT 1èreSong GabiPas encore d'évaluation
- Chapitre V ClonageDocument15 pagesChapitre V Clonageihcene aouesPas encore d'évaluation
- Glossaire GénétiqueDocument11 pagesGlossaire GénétiqueJojo75000Pas encore d'évaluation
- Génétique Et ÉpigénétiqueDocument41 pagesGénétique Et ÉpigénétiqueAdli Othmane100% (1)
- Chapitre 2 - Med 1 - Bio MolDocument12 pagesChapitre 2 - Med 1 - Bio Molyves100% (1)
- La Réplication de l'ADN - Fiche de Cours - SVT - SchoolMouv5 PDFDocument8 pagesLa Réplication de l'ADN - Fiche de Cours - SVT - SchoolMouv5 PDFjuini rawiaPas encore d'évaluation
- coursCorinneAbbadieM1bioinfopartie2 PDFDocument25 pagescoursCorinneAbbadieM1bioinfopartie2 PDFKhabtane AbdelhamidPas encore d'évaluation
- L'expression de L'information GénétiqueDocument15 pagesL'expression de L'information GénétiqueAMINE 16Pas encore d'évaluation
- COURS Abrg de Gnie Gntique Ver19Document29 pagesCOURS Abrg de Gnie Gntique Ver19awadiasmaPas encore d'évaluation
- Acides Nucléiques M4Document2 pagesAcides Nucléiques M4Doc AsmaPas encore d'évaluation
- Bilan Chapitre 2 La Replication de l' Adn Et Variabilite GenetiqueDocument3 pagesBilan Chapitre 2 La Replication de l' Adn Et Variabilite GenetiqueAymane JiraouiPas encore d'évaluation
- 5 - Construction Et CriblageDocument8 pages5 - Construction Et CriblageChibname ChibnamePas encore d'évaluation
- ED 2 de Biologie Cellulaire FMMDocument5 pagesED 2 de Biologie Cellulaire FMMbmnkhalilPas encore d'évaluation
- Compl Biologie Etudiant S-1Document43 pagesCompl Biologie Etudiant S-1aloys NdziePas encore d'évaluation
- Cours de Biotechnologies S5-DiapoDocument122 pagesCours de Biotechnologies S5-DiapoYahya RokniPas encore d'évaluation
- l12 Microbio Chap1Document9 pagesl12 Microbio Chap1Alhassane BahPas encore d'évaluation
- BIM Chapitre III Et IV PDFDocument37 pagesBIM Chapitre III Et IV PDFaya rezaiguiaPas encore d'évaluation
- Correction EMD TBBM (M1 Microbio, Mai 2023)Document6 pagesCorrection EMD TBBM (M1 Microbio, Mai 2023)biba.hiba625Pas encore d'évaluation
- Epreuve de GénétiqueDocument2 pagesEpreuve de GénétiqueIssouf BerthePas encore d'évaluation
- ABM5. Genetique BacterienneDocument17 pagesABM5. Genetique BacterienneMichel Tatelos NdeukamPas encore d'évaluation
- G02. Organisation Des Génomes (Cour)Document9 pagesG02. Organisation Des Génomes (Cour)marouaPas encore d'évaluation
- Chapitre I: L'expression de L'information GénétiqueDocument74 pagesChapitre I: L'expression de L'information Génétiqueahméd_facebPas encore d'évaluation
- Exem 1Document19 pagesExem 1Khaled KherrazPas encore d'évaluation
- EXPRESSION INFO GENETIQUE Prof OULAYADADocument37 pagesEXPRESSION INFO GENETIQUE Prof OULAYADAsamarasamounaPas encore d'évaluation
- V. Génome Et BiotechnologieDocument53 pagesV. Génome Et BiotechnologieLilouPas encore d'évaluation
- Biologie MoléculaireDocument10 pagesBiologie Moléculairemalak wxPas encore d'évaluation
- L'essentiel A Retenir Unité 2 (Partie 1 Et 2)Document6 pagesL'essentiel A Retenir Unité 2 (Partie 1 Et 2)rachidamine066Pas encore d'évaluation
- GENETIQUE FONDAMENTALE - Code Génétique - MoodleDocument4 pagesGENETIQUE FONDAMENTALE - Code Génétique - MoodleGh7Pas encore d'évaluation
- Début Chapitre 3 Expression Du Patrimoine Génétique 2023-2024Document3 pagesDébut Chapitre 3 Expression Du Patrimoine Génétique 2023-2024cloe.garrotPas encore d'évaluation
- Tout Sur La Réplication Du Matériel GénétiqueDocument10 pagesTout Sur La Réplication Du Matériel GénétiqueJéros Vigan85% (20)
- (Suite) GENETIQUE 1BACSMDocument11 pages(Suite) GENETIQUE 1BACSMnnadanada2901Pas encore d'évaluation
- 5éme Cours Génomique StructuraleDocument15 pages5éme Cours Génomique StructuraleHALIMAPas encore d'évaluation
- RemerciementsDocument3 pagesRemerciementssalama2011Pas encore d'évaluation
- 3) Objectif: Recueillir Les Perceptions À L'égard Des Lois Et Règlements EnvironnementauxDocument7 pages3) Objectif: Recueillir Les Perceptions À L'égard Des Lois Et Règlements Environnementauxsalama2011Pas encore d'évaluation
- Rapport de StageDocument3 pagesRapport de Stagesalama2011Pas encore d'évaluation
- Annexe 4 Document Envoyé Aux Experts Contexte Du ProjetDocument4 pagesAnnexe 4 Document Envoyé Aux Experts Contexte Du Projetsalama2011Pas encore d'évaluation
- 4 Présentation Des Résultats Et Discussion de L'enquête: 1. Pratiques Actuelles en Matière EnvironnementaleDocument13 pages4 Présentation Des Résultats Et Discussion de L'enquête: 1. Pratiques Actuelles en Matière Environnementalesalama2011Pas encore d'évaluation
- Outils de Genie Genetique - 5Document17 pagesOutils de Genie Genetique - 5salama2011Pas encore d'évaluation
- M1 PCPPDocument2 pagesM1 PCPPsalama2011Pas encore d'évaluation
- I-Caractéristiques Générales D'helicobacter Pylori: A - Présentation PhylogénétiqueDocument22 pagesI-Caractéristiques Générales D'helicobacter Pylori: A - Présentation Phylogénétiquesalama2011Pas encore d'évaluation
- Partie 2.1: Fonctionnement Et Utilisations Des Processus de FermentationDocument6 pagesPartie 2.1: Fonctionnement Et Utilisations Des Processus de Fermentationsalama2011Pas encore d'évaluation
- Partie 1.1: Les Bactéries: I) Définition Et Structure de La Cellule BactérienneDocument4 pagesPartie 1.1: Les Bactéries: I) Définition Et Structure de La Cellule Bactériennesalama2011Pas encore d'évaluation
- Bleu Du TrypanDocument69 pagesBleu Du TrypanMohamed ACHAHBAR100% (1)
- SVT 03 E1 2012 CorrigeDocument2 pagesSVT 03 E1 2012 Corrigefalilou loPas encore d'évaluation
- Voie Des Pentoses PhosphatesDocument26 pagesVoie Des Pentoses Phosphatescep100% (1)
- Af Jour 7 N 2Document1 pageAf Jour 7 N 2Zongo juliennePas encore d'évaluation
- DUTS2M6Document87 pagesDUTS2M6Steph 888Pas encore d'évaluation
- MaintDocument14 pagesMaintDib ZahiraPas encore d'évaluation
- Regulation D ADNDocument44 pagesRegulation D ADNYousra NanoPas encore d'évaluation
- L Essentiel de La Biologie Cellulaire 3 Ed Chapitre16Document18 pagesL Essentiel de La Biologie Cellulaire 3 Ed Chapitre16Vayo SonyPas encore d'évaluation
- Mécanisme de Défense Non SpécifiqueDocument4 pagesMécanisme de Défense Non SpécifiqueJoseph OnanaPas encore d'évaluation
- Fiche Mémorisation Chapitre 1 I) 2022Document3 pagesFiche Mémorisation Chapitre 1 I) 2022lafleche449Pas encore d'évaluation
- Fascicule GENETIQUEDocument48 pagesFascicule GENETIQUECho Min SooPas encore d'évaluation
- Micro Biolog I eDocument4 pagesMicro Biolog I eChescoPas encore d'évaluation
- BioMol Med2 Chap1-14 05 2022Document54 pagesBioMol Med2 Chap1-14 05 2022Denisco NkemgnePas encore d'évaluation
- Controle N°2 / Durée: 2h: Partie 1: Restitution Des Connaissances (5 Points)Document3 pagesControle N°2 / Durée: 2h: Partie 1: Restitution Des Connaissances (5 Points)nizarbouchnitaa100% (1)
- CoursDocument15 pagesCoursDaniel TeyouPas encore d'évaluation
- ExamDocument2 pagesExamRoukiyath AmoussaPas encore d'évaluation
- ReplicationDocument21 pagesReplicationMohamed Yassine MtarPas encore d'évaluation
- Hémostase Chirur DentaireDocument8 pagesHémostase Chirur DentaireTc TcPas encore d'évaluation
- TD 2Document3 pagesTD 2Yasmine GuerinikPas encore d'évaluation
- La Cellule EucaryoteDocument2 pagesLa Cellule EucaryoteIngrid Marchaux100% (1)
- TP1 ÉpithéliumsDocument56 pagesTP1 ÉpithéliumsHadrien De GreefPas encore d'évaluation
- Noyau InterphasiqueDocument29 pagesNoyau InterphasiqueOumarou KontaPas encore d'évaluation
- Bilan de L'unité 2Document7 pagesBilan de L'unité 2SALWA DAWDIPas encore d'évaluation
- Polycopie Du Cours Régulation Des Gènes Chez Les Procaryotes M1 Génétique MM GHARZOULIDocument64 pagesPolycopie Du Cours Régulation Des Gènes Chez Les Procaryotes M1 Génétique MM GHARZOULISarah SisiPas encore d'évaluation
- Histologie de La Peau 1315203247209 PDFDocument31 pagesHistologie de La Peau 1315203247209 PDFAhouri HamzaPas encore d'évaluation
- Physiologie de La Cellule Et de La MembraneDocument14 pagesPhysiologie de La Cellule Et de La MembraneKhaoula SahtouriPas encore d'évaluation
- Banque QCM CorrigésDocument22 pagesBanque QCM CorrigésTr LwPas encore d'évaluation
- 1 La Spermatogenese CoursDocument11 pages1 La Spermatogenese CoursNoahPas encore d'évaluation
- Cours2Basesde Données SpecialisésDocument49 pagesCours2Basesde Données SpecialiséslinePas encore d'évaluation
- Formulaire Gestion Donnees Comparaison InterLaboratoire HemogrammeDocument3 pagesFormulaire Gestion Donnees Comparaison InterLaboratoire HemogrammeOTOTEPas encore d'évaluation