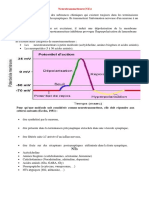
Vous aimerez peut-être aussi
- 1-CONVULSIONS CHEZ L'ENFANT Niveau M2 Sept 2018Document54 pages1-CONVULSIONS CHEZ L'ENFANT Niveau M2 Sept 2018dimitri messangaPas encore d'évaluation
- Inbound 2887955385113532655Document58 pagesInbound 2887955385113532655You SsefPas encore d'évaluation
- Cours Medoc SN 2017 + CommDocument203 pagesCours Medoc SN 2017 + CommBruce DO VANPas encore d'évaluation
- Mecanismes Neurobiologiques de L'epilepsieDocument16 pagesMecanismes Neurobiologiques de L'epilepsieMohamed Cisse75% (4)
- CAT Devant Les InsomniesDocument6 pagesCAT Devant Les InsomniesAsbat BaziPas encore d'évaluation
- CAT Devant Une Crise ÉpileptiqueDocument50 pagesCAT Devant Une Crise ÉpileptiqueIlhammoukhlissePas encore d'évaluation
- Cours Medoc SN 2015Document104 pagesCours Medoc SN 2015Bruce DO VANPas encore d'évaluation
- 18.antiépileptiques MedecineDocument51 pages18.antiépileptiques MedecineNi HelPas encore d'évaluation
- 03 - Etat de Mal ConvulsiveDocument4 pages03 - Etat de Mal ConvulsiveMedical studentPas encore d'évaluation
- PR K.yousfate Med1-Eme 2022Document33 pagesPR K.yousfate Med1-Eme 2022Hugekey PogchampPas encore d'évaluation
- AntiepileptiquesDocument24 pagesAntiepileptiquesAmiraBenhammouPas encore d'évaluation
- AntiepileptiquesDocument56 pagesAntiepileptiquesAnnab TakiPas encore d'évaluation
- Les Nouvelles Stratégies Thérapeutiques Ciblant La Maladie D'alzheimer FFDocument5 pagesLes Nouvelles Stratégies Thérapeutiques Ciblant La Maladie D'alzheimer FFabelarbi290Pas encore d'évaluation
- ANTIEPILEPTIQUESDocument3 pagesANTIEPILEPTIQUESNihad NshPas encore d'évaluation
- 4 - Les Épilepsies 2 Eme PartieDocument8 pages4 - Les Épilepsies 2 Eme Partiesally crowePas encore d'évaluation
- 1 PHARMACOLOGIE ANTIEPILEPTIQUES SupportDocument9 pages1 PHARMACOLOGIE ANTIEPILEPTIQUES SupportFadilouPas encore d'évaluation
- Prise en Charge D'une Crise ConvulsiveDocument17 pagesPrise en Charge D'une Crise Convulsivesamir hacibPas encore d'évaluation
- Neuro PM - New1Document58 pagesNeuro PM - New1Louiza HouatPas encore d'évaluation
- Sémiologie Des Crises Épileptiques-FinaleDocument4 pagesSémiologie Des Crises Épileptiques-Finaleoumaimahenteti5Pas encore d'évaluation
- SEPI 8 Et 9Document17 pagesSEPI 8 Et 9leybadaraccoPas encore d'évaluation
- RP 176 EPU IsapofDocument3 pagesRP 176 EPU Isapofsara laadamiPas encore d'évaluation
- 13-Convulsion de L'enfantDocument6 pages13-Convulsion de L'enfantAhmed HoussemPas encore d'évaluation
- EpilepsieDocument25 pagesEpilepsietemaPas encore d'évaluation
- Medicaments Antileptiques (Full Permission)Document33 pagesMedicaments Antileptiques (Full Permission)Belkacem HantaliPas encore d'évaluation
- Cours 2 PharmacoDocument25 pagesCours 2 PharmacoNOUR YOUSSEFPas encore d'évaluation
- 26-Epilepsie Cours Commun 2020 7Document33 pages26-Epilepsie Cours Commun 2020 7Sahar ChnitirPas encore d'évaluation
- Sibon EpilepsiesDocument66 pagesSibon Epilepsiesgustel vinnyPas encore d'évaluation
- ConvulsionsDocument20 pagesConvulsionsBouhannache MeryemPas encore d'évaluation
- 4 e SEQ TD 2021 - LBBPP PDFDocument5 pages4 e SEQ TD 2021 - LBBPP PDFjames taboulaPas encore d'évaluation
- Convulsion Etat de Mal ConvulsifDocument6 pagesConvulsion Etat de Mal Convulsifkaoutar blcPas encore d'évaluation
- Bac S 2018 - Amérique Du Sud - SVT - Académie de BesançonDocument10 pagesBac S 2018 - Amérique Du Sud - SVT - Académie de Besançongzqg9cp7mtPas encore d'évaluation
- Cours Epilepsie Sommeil Mars 2012 VHNMDocument47 pagesCours Epilepsie Sommeil Mars 2012 VHNMMihaela-Alexandra PopPas encore d'évaluation
- Les Epilepsies Cour ChirdentDocument5 pagesLes Epilepsies Cour Chirdentbouchemlabilal95Pas encore d'évaluation
- 7-Système Nerveux Végétatif-1Document17 pages7-Système Nerveux Végétatif-1fancinadounosiamariePas encore d'évaluation
- CF24Document2 pagesCF24George LupuPas encore d'évaluation
- NeurotropesDocument44 pagesNeurotropesBENCHABANE OUSSAMAPas encore d'évaluation
- NEUROMEDIATEURSDocument40 pagesNEUROMEDIATEURSBENCHABANE OUSSAMAPas encore d'évaluation
- SF1 CNSDocument63 pagesSF1 CNSdjuroPas encore d'évaluation
- 7antalgiques Et SalycilésDocument6 pages7antalgiques Et SalycilésBel KisPas encore d'évaluation
- Neuro 2Document12 pagesNeuro 2tinaPas encore d'évaluation
- 7 EpilepsiesDocument12 pages7 EpilepsiesGoogle PlzPas encore d'évaluation
- Em5convulsion de L'enfant-1Document31 pagesEm5convulsion de L'enfant-1Hamza BIKIPas encore d'évaluation
- 4 - Épilepsies Cour 2021Document69 pages4 - Épilepsies Cour 2021fatima zahra banhssinPas encore d'évaluation
- 4 - ÉpilepsiesDocument11 pages4 - ÉpilepsieshouriaPas encore d'évaluation
- Anesthesiques CoursDocument37 pagesAnesthesiques CoursAlexandre Kpangny Béni100% (1)
- Crises Convulsives Pour M2 2017Document54 pagesCrises Convulsives Pour M2 2017Nkuh Cliford NgangePas encore d'évaluation
- Les Neuromediateurs 2024Document94 pagesLes Neuromediateurs 2024Corona CoronaPas encore d'évaluation
- Les NeurotransmetteursDocument15 pagesLes NeurotransmetteursNo Comment.Pas encore d'évaluation
- Cours Neuro-PharmaDocument4 pagesCours Neuro-PharmaDan KAYEMBEPas encore d'évaluation
- EpilepsieDocument11 pagesEpilepsieSamar OmraniPas encore d'évaluation
- INSULINE ET SES EFFETS PHYSIOLOGIQUESDocument6 pagesINSULINE ET SES EFFETS PHYSIOLOGIQUESMariettePas encore d'évaluation
- Sémiologie Des Crises 2020Document9 pagesSémiologie Des Crises 2020Mehdi BayanaPas encore d'évaluation
- Les Médicaments PsychotropesDocument8 pagesLes Médicaments PsychotropesJaşmĭnę Bēn AmởrPas encore d'évaluation
- Veille SommeilDocument8 pagesVeille Sommeilkhouloud djebabraPas encore d'évaluation
- ÉpilepsieDocument6 pagesÉpilepsieSouheil MansouriPas encore d'évaluation
- Biologie de La Dépression (28.03)Document10 pagesBiologie de La Dépression (28.03)oketarpeqPas encore d'évaluation
- 1 - Epilepsie Et Crise ConvulsiveDocument24 pages1 - Epilepsie Et Crise ConvulsiveSee DownPas encore d'évaluation
- Médicaments Du S.parasympatiqueDocument23 pagesMédicaments Du S.parasympatiqueDemba Tahirou DIOPPas encore d'évaluation
- Lignelet Medicaments AntileptiquesDocument33 pagesLignelet Medicaments AntileptiquesMina KDPas encore d'évaluation
- BIOLE102 Neurobiologie: LégendeDocument117 pagesBIOLE102 Neurobiologie: LégendeDaniela RiosPas encore d'évaluation
- Physiologie Cours COMPLETDocument492 pagesPhysiologie Cours COMPLETProjojo99Pas encore d'évaluation
- Le Tissu Nerveux (Mode de Compatibilité)Document64 pagesLe Tissu Nerveux (Mode de Compatibilité)عبد السميع بن أحمدPas encore d'évaluation
- Résumé G.Chakib NerveuxDocument5 pagesRésumé G.Chakib NerveuxMaroua AbzPas encore d'évaluation
- Pensez-Vous Que La Théorie Cellulaire Telle Qu'elle Est Définie Est Généralisable À Tout Le Vivant?Document2 pagesPensez-Vous Que La Théorie Cellulaire Telle Qu'elle Est Définie Est Généralisable À Tout Le Vivant?Nathan PululuPas encore d'évaluation
- Tissus NerveuxDocument19 pagesTissus NerveuxGuenzet Akila100% (1)
- Neurophysiologie Et NeuroanatomieDocument141 pagesNeurophysiologie Et NeuroanatomieDuboisPas encore d'évaluation
- Le Tissu NerveuxDocument16 pagesLe Tissu NerveuxZizou HamdiPas encore d'évaluation
- Tissu NerveuxDocument36 pagesTissu Nerveuxfederica.lamaPas encore d'évaluation
- Organisation Du Système NerveuxDocument19 pagesOrganisation Du Système NerveuxMaelle DURANDPas encore d'évaluation
- Physiopathologie NeuroscienceDocument46 pagesPhysiopathologie NeuroscienceYas MiiinaaPas encore d'évaluation
- Andre Et Al 1999Document145 pagesAndre Et Al 1999Eduardo AnguloPas encore d'évaluation
- Cours Physiologie Animale 1-1Document150 pagesCours Physiologie Animale 1-1Mina OussarPas encore d'évaluation
- Met-302459-Encephalopathie Hepatique de La Physiopathologie A La Prise en Charge Therapeutique - Wr3bGn8AAQEAAGZkbyMAAAAE-ADocument7 pagesMet-302459-Encephalopathie Hepatique de La Physiopathologie A La Prise en Charge Therapeutique - Wr3bGn8AAQEAAGZkbyMAAAAE-ANoadia RafararisonPas encore d'évaluation
- Le Programme Pour Bien Nourrir Votre Cerveau 2019 Jean Marie BourreDocument367 pagesLe Programme Pour Bien Nourrir Votre Cerveau 2019 Jean Marie BourreMoussaPas encore d'évaluation
- 3 - Œdème Cérébral Nouvelles Pistes ThérapeutiquesDocument8 pages3 - Œdème Cérébral Nouvelles Pistes ThérapeutiquesyaalaPas encore d'évaluation
- CN1 Neurophysiologie A. Organisation Du Système Nerveux: Comment Peut-On Expliquer Une Telle Évolution?Document23 pagesCN1 Neurophysiologie A. Organisation Du Système Nerveux: Comment Peut-On Expliquer Une Telle Évolution?Oriane plcnPas encore d'évaluation
- Introduction NeurophysiologieDocument8 pagesIntroduction NeurophysiologieKatia ZakPas encore d'évaluation
- Fiche Métabolisme SNC Et GR Organe Gluco DependantsDocument6 pagesFiche Métabolisme SNC Et GR Organe Gluco DependantsNour klPas encore d'évaluation
- BIO 270 Physiologie Cellulaire Générale 2020 PDFDocument125 pagesBIO 270 Physiologie Cellulaire Générale 2020 PDFTONDI ABDOU CHACOURPas encore d'évaluation
- Neurophysiologie 2Document49 pagesNeurophysiologie 2123kiwiPas encore d'évaluation
- CM 1 Neurophysiologie - Introduction Et Cellules Gliales N GuéroutDocument82 pagesCM 1 Neurophysiologie - Introduction Et Cellules Gliales N GuéroutfezbejfPas encore d'évaluation
- Tissus NerveuxDocument106 pagesTissus Nerveuxval35000Pas encore d'évaluation
- Veurosciences Cognitives, Les - Nicole Fiori-Duharcourt & Frederic Isel PDFDocument104 pagesVeurosciences Cognitives, Les - Nicole Fiori-Duharcourt & Frederic Isel PDFMourad RoudjPas encore d'évaluation
- Physiologie Humaine Chap 2: Bases Cellulaires Du Contrôle NerveuxDocument77 pagesPhysiologie Humaine Chap 2: Bases Cellulaires Du Contrôle NerveuxLaurinePas encore d'évaluation
- Cours Neurohisto PCEM1 2006-2007Document50 pagesCours Neurohisto PCEM1 2006-2007Wiwi LauraPas encore d'évaluation
- SYSTEME NERVEUX RechDocument14 pagesSYSTEME NERVEUX Rechariane.vasseur29Pas encore d'évaluation
- 1e0d9bb63f DDocument27 pages1e0d9bb63f DsahexePas encore d'évaluation
- NeurophysiologieDocument13 pagesNeurophysiologieCath Lemeunier100% (1)
- Chapitre 2 M Canismes Et Notions L Mentai 2020 Epilepsies de Lenfant deDocument10 pagesChapitre 2 M Canismes Et Notions L Mentai 2020 Epilepsies de Lenfant deAtarki MeriemPas encore d'évaluation