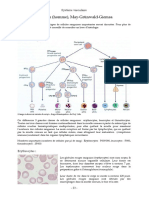
Vous aimerez peut-être aussi
- 4 PlaquettesDocument4 pages4 PlaquettessafemindPas encore d'évaluation
- CivdDocument8 pagesCivdArdeleanu CostinPas encore d'évaluation
- Discordances Dans Les Groupages Sanguins AboDocument16 pagesDiscordances Dans Les Groupages Sanguins AboLandry TengaPas encore d'évaluation
- Exploration de L'hémostaseDocument18 pagesExploration de L'hémostaseKhaoula SahtouriPas encore d'évaluation
- Hemostase New1Document61 pagesHemostase New1Dominique Ilunga mbambiPas encore d'évaluation
- Hemato HémostaseDocument17 pagesHemato HémostaseN.Pas encore d'évaluation
- Physiologie Et Sémiologie de L'hémostaseDocument47 pagesPhysiologie Et Sémiologie de L'hémostasesafemind100% (2)
- Physiologie de L - Hã©mostase - 2004 - EMC - DentisterieDocument11 pagesPhysiologie de L - Hã©mostase - 2004 - EMC - Dentisterieamira benhamoudaPas encore d'évaluation
- Physiologie Et Exploration Del Hémostase-Poly P2-2011Document40 pagesPhysiologie Et Exploration Del Hémostase-Poly P2-2011Nebel GesangPas encore d'évaluation
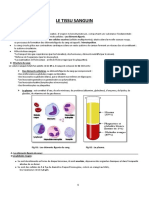
- III - 4 Le Tissu SanguinDocument14 pagesIII - 4 Le Tissu SanguinalmnaouarPas encore d'évaluation
- HémostaseDocument62 pagesHémostaseAnas SabbanePas encore d'évaluation
- 1.PHYSIOLOGIE HEMOSTASE PRIMAIREDocument3 pages1.PHYSIOLOGIE HEMOSTASE PRIMAIREliliwoody986Pas encore d'évaluation
- Hemato4an - Physiologie Hemostase CoursDocument5 pagesHemato4an - Physiologie Hemostase Coursbr manelPas encore d'évaluation
- 175 Traitement AntithrombotiqueDocument37 pages175 Traitement AntithrombotiqueYahia BenaliPas encore d'évaluation
- Tissu SanguinDocument25 pagesTissu Sanguingroupe_svi3100% (1)
- Chapitre 1 SangDocument43 pagesChapitre 1 SangNorhaine Nina100% (1)
- Physiologie Et Exploration de LDocument8 pagesPhysiologie Et Exploration de LAymenPas encore d'évaluation
- Hemostase-1Document10 pagesHemostase-1MB SKOFIELDPas encore d'évaluation
- Cours Physiologie de L'hémostase Et Méthodes D'explorationDocument172 pagesCours Physiologie de L'hémostase Et Méthodes D'explorationHKIMI KhouloudPas encore d'évaluation
- Copie de Chap 3) L'hémostaseDocument7 pagesCopie de Chap 3) L'hémostaseNT “NitixProd” PRODPas encore d'évaluation
- HemostaseDocument8 pagesHemostaseHanane JibetPas encore d'évaluation
- TSG 2.11Document40 pagesTSG 2.11Adonis SerghiniPas encore d'évaluation
- Tissu Sanguin Pearl ComlanDocument58 pagesTissu Sanguin Pearl Comlansehoemmanuel15Pas encore d'évaluation
- Oral Cours Physiopath ArterioscleroseDocument14 pagesOral Cours Physiopath Arteriosclerosemalick SemourPas encore d'évaluation
- Tissu SanguinDocument15 pagesTissu SanguinAlaa Sobhi100% (1)
- 1.hémostase - Physiologie, Explorations Et Interprétation CliniqueDocument17 pages1.hémostase - Physiologie, Explorations Et Interprétation CliniqueNaruto erdPas encore d'évaluation
- LDocument4 pagesLMa RyPas encore d'évaluation
- Physiologie de L'hémostase PrimaireDocument5 pagesPhysiologie de L'hémostase PrimaireI.m. Daniel100% (1)
- 04 Systeme VasculaireDocument9 pages04 Systeme VasculaireSamir MeddouraPas encore d'évaluation
- Hémostase Chirur DentaireDocument8 pagesHémostase Chirur DentaireTc TcPas encore d'évaluation
- HEMOSTASE PRIMAIRE, 2ndaire, FibrynolyseDocument94 pagesHEMOSTASE PRIMAIRE, 2ndaire, FibrynolyseNawal Ibrahim Ahmed SaidPas encore d'évaluation
- HemostaseDocument26 pagesHemostaseLeon ChipondaPas encore d'évaluation
- 3 HematopoieseDocument12 pages3 HematopoieseBalayira BakaryPas encore d'évaluation
- A9 10 11 Globul Roug Blanc PlakDocument8 pagesA9 10 11 Globul Roug Blanc PlakFernando KollaPas encore d'évaluation
- Physiologie Animale Cours 2Document53 pagesPhysiologie Animale Cours 2g69wwpfw5kPas encore d'évaluation
- Le SangDocument46 pagesLe SangJoronavalona RASAMIMANANAPas encore d'évaluation
- Hemostase Primaire 2018Document26 pagesHemostase Primaire 2018fifi fifiPas encore d'évaluation
- AthéromeDocument17 pagesAthéromeFreddy CarelPas encore d'évaluation
- 0.INTRODUCTION A L’HEMOSTASEDocument2 pages0.INTRODUCTION A L’HEMOSTASEliliwoody986Pas encore d'évaluation
- 6 - HematologieDocument36 pages6 - HematologieSamir AggounePas encore d'évaluation
- 34 Plaquettes SanguinesDocument8 pages34 Plaquettes SanguinesmaryPas encore d'évaluation
- 2 Cours 3-4 Physiologie Des Grandes Fonctions - PPT NDocument73 pages2 Cours 3-4 Physiologie Des Grandes Fonctions - PPT NIlyeSePas encore d'évaluation
- Atherome Epidemio Et Physiopath PDFDocument7 pagesAtherome Epidemio Et Physiopath PDFyouPas encore d'évaluation
- I-9-128-Athérome Épidémiologie, PhysiopathologieDocument7 pagesI-9-128-Athérome Épidémiologie, PhysiopathologieTaxo HaPas encore d'évaluation
- Universite de LubumbashiDocument5 pagesUniversite de Lubumbashiktk9gnvqjhPas encore d'évaluation
- Elements D'hemostaseDocument10 pagesElements D'hemostasegjobPas encore d'évaluation
- Mega Cary Poi EseDocument33 pagesMega Cary Poi EseTaoufik Ben HoumichPas encore d'évaluation
- Polynucléaires NeutrophilesDocument9 pagesPolynucléaires Neutrophileshhaammiidd11994499Pas encore d'évaluation
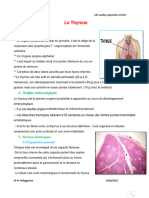
- Le ThymusDocument7 pagesLe ThymusManella Abdel DjebarPas encore d'évaluation
- Tissu SanguinDocument8 pagesTissu Sanguintim SssPas encore d'évaluation
- Le Tissu ConjonctifDocument55 pagesLe Tissu ConjonctifLisa FUzeliePas encore d'évaluation
- Histo EndoDocument4 pagesHisto EndochadelkettPas encore d'évaluation
- HématopoïèseDocument10 pagesHématopoïèseFayad BouraimaPas encore d'évaluation
- 39 ch33 356 368 9782294755163 CopieDocument9 pages39 ch33 356 368 9782294755163 CopieYahia BenaliPas encore d'évaluation
- 13Document12 pages13Dan BiPas encore d'évaluation
- Physiologie de L'hémostaseDocument5 pagesPhysiologie de L'hémostaseNouna nouniaPas encore d'évaluation
- Physiologie de L'hémostase: Par Françoise BalédentDocument6 pagesPhysiologie de L'hémostase: Par Françoise BalédentMohammad Al-AneedPas encore d'évaluation
- Surrénales: Les Grands Articles d'UniversalisD'EverandSurrénales: Les Grands Articles d'UniversalisPas encore d'évaluation
- Membranes cellulaires: Les Grands Articles d'UniversalisD'EverandMembranes cellulaires: Les Grands Articles d'UniversalisPas encore d'évaluation
- Tissu ConjonctifDocument11 pagesTissu Conjonctifgerry iPas encore d'évaluation
- Tissu Musculaire Strié SquelettiqueDocument10 pagesTissu Musculaire Strié Squelettiquegerry iPas encore d'évaluation
- TM LisseDocument7 pagesTM Lissegerry iPas encore d'évaluation
- Tableau Des Hiragana VERSION MANGA - Comment L'utiliserDocument2 pagesTableau Des Hiragana VERSION MANGA - Comment L'utiliserHarperPas encore d'évaluation
- Fiche Pratique Comprendre Les Resultats Des Analyses de SangDocument2 pagesFiche Pratique Comprendre Les Resultats Des Analyses de SangFaiza RebbouhPas encore d'évaluation
- Damage Control2017Document12 pagesDamage Control2017ElbordjiPas encore d'évaluation
- TD12 TransfusionDocument22 pagesTD12 TransfusionAbderrahmenPas encore d'évaluation
- Diagnostic D'une Anémie - 2016 - JaccardDocument65 pagesDiagnostic D'une Anémie - 2016 - JaccardHmed Ben MohamedPas encore d'évaluation
- Rapport de StageDocument22 pagesRapport de StagenonoliliaPas encore d'évaluation
- Présentation SysmexDocument18 pagesPrésentation SysmexJu Ju100% (1)
- Utilisation Du RFVIIA Novoseven Lors DunDocument4 pagesUtilisation Du RFVIIA Novoseven Lors DunAlain SoucotPas encore d'évaluation
- Cicatrisation en Milieu Buccale PDFDocument218 pagesCicatrisation en Milieu Buccale PDFSmailPas encore d'évaluation
- Hémostase Primaire (Retapé)Document5 pagesHémostase Primaire (Retapé)Chahra ZedPas encore d'évaluation
- Le Manuel Du Généraliste Les Anomalies Biologiques Courantes 2017Document145 pagesLe Manuel Du Généraliste Les Anomalies Biologiques Courantes 2017lotfi cyclistePas encore d'évaluation
- Guide Des Dons Métropole 2021Document17 pagesGuide Des Dons Métropole 2021seydinay3Pas encore d'évaluation
- Anemies de L'enfantDocument46 pagesAnemies de L'enfantDOCTOR HOUSEPas encore d'évaluation
- Qe Amrani IlyothyDocument16 pagesQe Amrani IlyothyOUMAIMA EL CHAALPas encore d'évaluation
- 5 Valla Place Des Anticoagulants Au Cours de La CirrhoseDocument35 pages5 Valla Place Des Anticoagulants Au Cours de La CirrhoseIrina GîrleanuPas encore d'évaluation
- Hémogramme Numeration Des Erythrocytes Et Dosage de L'hemoglobineDocument6 pagesHémogramme Numeration Des Erythrocytes Et Dosage de L'hemoglobinebouabidPas encore d'évaluation
- 1 s2.0 S0515370014002651 MainDocument3 pages1 s2.0 S0515370014002651 MainSniper4DzPas encore d'évaluation
- Anémie Hémolytique Autoimmune (AHAI) : Guide Des Analyses SpécialiséesDocument3 pagesAnémie Hémolytique Autoimmune (AHAI) : Guide Des Analyses SpécialiséesIssa CharkawiPas encore d'évaluation
- Mondet Julie p02Document19 pagesMondet Julie p02Desthin Mounzéo NgomaPas encore d'évaluation
- Tableau CPNDocument1 pageTableau CPNElvira Cortes JaramilloPas encore d'évaluation
- Facteur VDocument2 pagesFacteur Vvilaxax669Pas encore d'évaluation
- Info ProduitDocument1 pageInfo ProduitClaudia DiţaPas encore d'évaluation
- TP Difficultés de Groupage SanguinDocument39 pagesTP Difficultés de Groupage SanguinLamia Lalam33% (3)
- Les AnemiesDocument20 pagesLes Anemiesfatoutraore2345Pas encore d'évaluation
- QCM Concours Blanc 2010 2011 1Document8 pagesQCM Concours Blanc 2010 2011 1Aicha GrPas encore d'évaluation
- HemostaseDocument20 pagesHemostaseJessica ZoghbyPas encore d'évaluation
- Listing Global ChristianDocument3 pagesListing Global Christianchristian leubouPas encore d'évaluation
- Envois Exterieurs: 2300087846 Bocobza CecileDocument4 pagesEnvois Exterieurs: 2300087846 Bocobza CecilececilejoulinPas encore d'évaluation
- Les Anémies Arégénératives - 1Document73 pagesLes Anémies Arégénératives - 1AbdelOuahidSenhadjiPas encore d'évaluation