Vous aimerez peut-être aussi
- Génétique: Les Grands Articles d'UniversalisD'EverandGénétique: Les Grands Articles d'UniversalisPas encore d'évaluation
- Cours 9 MutationsDocument3 pagesCours 9 Mutationsdjassa djassaPas encore d'évaluation
- L'infertilité N'est Pas Seulement Un Problème Du CoupleD'EverandL'infertilité N'est Pas Seulement Un Problème Du CouplePas encore d'évaluation
- Mutations Genetique JRDocument3 pagesMutations Genetique JRJean pie jean piePas encore d'évaluation
- Mutation Génomique Mesmin - 2Document16 pagesMutation Génomique Mesmin - 2Marius Ganhouegnon100% (5)
- Mecanismes de Reparation de L'adn Chez Les PocaryotesDocument17 pagesMecanismes de Reparation de L'adn Chez Les PocaryotesMeguerouni GhaniaPas encore d'évaluation
- Génétique - Les MutationsDocument4 pagesGénétique - Les MutationsThomas GiglaPas encore d'évaluation
- Techniques D'étude Du Génome HumainDocument21 pagesTechniques D'étude Du Génome Humainkhediri hibaPas encore d'évaluation
- Chap 2 MutationsDocument20 pagesChap 2 MutationsHassanPas encore d'évaluation
- 8a p2 Ue1 Cours 4 Du Gène À La Protéine Mutante Partie1 RonéoDocument14 pages8a p2 Ue1 Cours 4 Du Gène À La Protéine Mutante Partie1 RonéoksllmkPas encore d'évaluation
- Chapitre 4 Les MutationsDocument12 pagesChapitre 4 Les Mutationsoussamabennaceur2000Pas encore d'évaluation
- SciencesDocument1 pageSciencesmax.demoerloosePas encore d'évaluation
- MutationsDocument61 pagesMutationslobnaPas encore d'évaluation
- Mutation Et Mécanismes de Réparation de lADNDocument57 pagesMutation Et Mécanismes de Réparation de lADNMichel Tatelos NdeukamPas encore d'évaluation
- Microbio s3 Cours 8Document66 pagesMicrobio s3 Cours 8Jamila ridaPas encore d'évaluation
- Stabilite-et-Variabilite-des-GenomesDocument6 pagesStabilite-et-Variabilite-des-Genomesimenbmc1Pas encore d'évaluation
- Génotoxicité Mutagénèse TératogenèseDocument10 pagesGénotoxicité Mutagénèse TératogenèseRaid YaSsinePas encore d'évaluation
- Chapitre 3 Les Événements Génétiques DaleDocument15 pagesChapitre 3 Les Événements Génétiques Dalemanalchannel5Pas encore d'évaluation
- Biotech-Master I - Ufhb-2013 Chap 2fDocument53 pagesBiotech-Master I - Ufhb-2013 Chap 2fjeanlouiskouakouPas encore d'évaluation
- MutationDocument5 pagesMutationredouanePas encore d'évaluation
- Mutation Et Mécanismes de Réparation de LDocument9 pagesMutation Et Mécanismes de Réparation de LFati HaPas encore d'évaluation
- CHAPITRE Mecanisme de L'évolution-ConvertiDocument3 pagesCHAPITRE Mecanisme de L'évolution-ConvertiSoumana SogobaPas encore d'évaluation
- Ogm Et Mutations Sabine MehoumeDocument15 pagesOgm Et Mutations Sabine MehoumeMarius Ganhouegnon100% (1)
- PlasmideDocument35 pagesPlasmidea.itatahinePas encore d'évaluation
- cours-genotoxicologie2-2Document218 pagescours-genotoxicologie2-2Nourben MuchyaPas encore d'évaluation
- Autre Cours Genetique Mutation PDFDocument8 pagesAutre Cours Genetique Mutation PDFlobnaPas encore d'évaluation
- Les Mutations Et La Génétique Des PopulationsDocument23 pagesLes Mutations Et La Génétique Des PopulationsFaiza HPas encore d'évaluation
- Cours 8 Les MutationsDocument8 pagesCours 8 Les MutationsFarah B. BtoushPas encore d'évaluation
- Chapitre 3 . PRINCIPAUX EFFETS PHYSIOTOXICOLOGIQUESDocument11 pagesChapitre 3 . PRINCIPAUX EFFETS PHYSIOTOXICOLOGIQUESahmed.213karPas encore d'évaluation
- Theme 1 Chap 4 GenetiqueDocument4 pagesTheme 1 Chap 4 GenetiquejeanlouiskouakouPas encore d'évaluation
- Activité 7 Version 2024Document2 pagesActivité 7 Version 2024Ahmedrayane NamlyPas encore d'évaluation
- Thème 1, Chap3 Varaibilité Genetique Et MutationDocument3 pagesThème 1, Chap3 Varaibilité Genetique Et MutationThe CelestinyPas encore d'évaluation
- Tpe Biologie Le Bon - 102549Document19 pagesTpe Biologie Le Bon - 102549ismaeilndjiana26Pas encore d'évaluation
- Cours MMI Intro-Transferts 2021 2Document14 pagesCours MMI Intro-Transferts 2021 2Rozalina rania SallamiPas encore d'évaluation
- Innovations Génétiques - WikipédiaDocument23 pagesInnovations Génétiques - WikipédiaKaton dit abdramane CoulibalyPas encore d'évaluation
- Get MutDocument122 pagesGet MutAbdu LazizPas encore d'évaluation
- 5 .Genetique BacterienneDocument8 pages5 .Genetique BacterienneIliass NaouadirPas encore d'évaluation
- Diversité M1 BPODocument24 pagesDiversité M1 BPOlli510265Pas encore d'évaluation
- Cours de GenetiqueDocument33 pagesCours de Genetiquebonneaug8Pas encore d'évaluation
- Mutation de l'ADNDocument6 pagesMutation de l'ADNZakaria FerkachePas encore d'évaluation
- 15 - Mécanisme de La CarcinogénèseDocument49 pages15 - Mécanisme de La CarcinogénèseManel Bouarbi100% (1)
- Tâches Complexes de SVTDocument2 pagesTâches Complexes de SVTMaeva Costa BeziatPas encore d'évaluation
- Stabilité Et Variabilité Des Génômes Et ÉvolutionDocument6 pagesStabilité Et Variabilité Des Génômes Et ÉvolutionHellie WrightPas encore d'évaluation
- Cancerogenese Et Croissance Tumorale 231010 165953Document6 pagesCancerogenese Et Croissance Tumorale 231010 165953moussaoued493Pas encore d'évaluation
- 1spé T1 Chapitre3 CoursDocument8 pages1spé T1 Chapitre3 Coursabdirahman abdillahiPas encore d'évaluation
- TermS PARTIE B Chapitre 3Document27 pagesTermS PARTIE B Chapitre 3sandra.le.duPas encore d'évaluation
- EpigenetiqueDocument4 pagesEpigenetiqueLyes BazaPas encore d'évaluation
- BIO OGM PrésentationDocument4 pagesBIO OGM PrésentationYoram CaroyPas encore d'évaluation
- UE Endocrinologie Cellulaire M2PANDocument13 pagesUE Endocrinologie Cellulaire M2PANteresa ekouerembaPas encore d'évaluation
- GénotoxicitéDocument5 pagesGénotoxicitéIkram HamachePas encore d'évaluation
- CoursDocument16 pagesCoursKoki kokiPas encore d'évaluation
- Bac S 2018 EAT CorrigéDocument4 pagesBac S 2018 EAT CorrigéLETUDIANTPas encore d'évaluation
- Chapitre 3 Recombinaison GénétiqueDocument5 pagesChapitre 3 Recombinaison GénétiqueAbboud DjamaaPas encore d'évaluation
- Student PHYTOHORMONE 2021 2022Document46 pagesStudent PHYTOHORMONE 2021 2022IDTH EBM ApplicationPas encore d'évaluation
- SVT Sujets Type 1 (À Copier)Document4 pagesSVT Sujets Type 1 (À Copier)cristqlinePas encore d'évaluation
- Undone 3Document24 pagesUndone 3rdaPas encore d'évaluation
- Chapitre 10 EmbryologieA1 Tératogénèse Djeffal PDFDocument7 pagesChapitre 10 EmbryologieA1 Tératogénèse Djeffal PDFVéto SamPas encore d'évaluation
- Microbiologie II-SV5: Pr. Nadia El KadmiriDocument146 pagesMicrobiologie II-SV5: Pr. Nadia El KadmiriHamidPas encore d'évaluation
- ChapIV Signalisation Et RégulationDocument37 pagesChapIV Signalisation Et RégulationMamadou lamine DiattaPas encore d'évaluation
- Relation Géne Phénotype EnvironnementDocument3 pagesRelation Géne Phénotype Environnementle sage100% (1)
- Matière Microorganismes PhotosynthétiquesDocument114 pagesMatière Microorganismes Photosynthétiqueshayet GhalemPas encore d'évaluation
- Chap 3Document2 pagesChap 3Zineb ThiyfaPas encore d'évaluation
- Cours de Virologie Systematique Fss Ul Ls5Document85 pagesCours de Virologie Systematique Fss Ul Ls5BamonPas encore d'évaluation
- Synthèse Des Biomolécules M2 Biochimie 2020 - 2021Document29 pagesSynthèse Des Biomolécules M2 Biochimie 2020 - 2021Aissa TlbPas encore d'évaluation
- Morphologie Des BacteriesDocument26 pagesMorphologie Des Bacteriesalmnaouar100% (1)
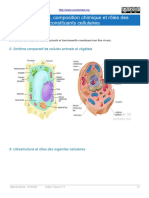
- Ultrastructure Composition Chimique Et Roles de Constituants CellulaireDocument4 pagesUltrastructure Composition Chimique Et Roles de Constituants Cellulairefyvsgbj92bPas encore d'évaluation
- Génétique Bactérienne Les Échanges Génétiques, Évolution, Plasticité Des Génomes Bactériens PDFDocument13 pagesGénétique Bactérienne Les Échanges Génétiques, Évolution, Plasticité Des Génomes Bactériens PDFkerkour-abd1523Pas encore d'évaluation
- Devoir Corrigé de Contrôle N°3 - SVT - 1ère AS (2014-2015) MR Mzid MouradDocument2 pagesDevoir Corrigé de Contrôle N°3 - SVT - 1ère AS (2014-2015) MR Mzid MouradChokri Besbes100% (1)
- Cellules - Metabolisme Cellules - Photosynthese - Respiration - Seconde SVT 2023Document9 pagesCellules - Metabolisme Cellules - Photosynthese - Respiration - Seconde SVT 2023tototomato 17Pas encore d'évaluation
- BiologieDocument14 pagesBiologieFéz EyPas encore d'évaluation
- Généralités Sur La CelluleDocument14 pagesGénéralités Sur La CelluleJulien MinotPas encore d'évaluation
- AFADocument4 pagesAFADodji Kekeli Eben-EzerPas encore d'évaluation
- SVT MoDocument3 pagesSVT MoFrancesca GermainPas encore d'évaluation
- L'essentiel de La Biologie Cellulaire: Rappels de Cours Et QCM CorrigésDocument15 pagesL'essentiel de La Biologie Cellulaire: Rappels de Cours Et QCM CorrigésAKEMPas encore d'évaluation
- Exp Sur Le ChatDocument32 pagesExp Sur Le Chatel_guitar_istPas encore d'évaluation
- BioQ12018 PDFDocument36 pagesBioQ12018 PDFzineb EZZINEPas encore d'évaluation
- Cellule Bacterienne Chap II 2 PartieDocument26 pagesCellule Bacterienne Chap II 2 Partieهشام شريفيPas encore d'évaluation
- 1ère S2 D1 S.V.T SEM1 021Document2 pages1ère S2 D1 S.V.T SEM1 021Papa Adama Diakhate100% (3)
- À La Découverte D Un Milieu NaturelDocument5 pagesÀ La Découverte D Un Milieu Naturelal mounir alaouiPas encore d'évaluation
- Exposé de Génie Génétique-ConvertiDocument9 pagesExposé de Génie Génétique-ConvertiAhcene BouflighaPas encore d'évaluation
- UE2 La-Membrane-PlasmiqueDocument17 pagesUE2 La-Membrane-Plasmiquesun-nee-chan9Pas encore d'évaluation
- 3eme - Thème: La Planète Terre, L'environnement Et L'action HumaineDocument16 pages3eme - Thème: La Planète Terre, L'environnement Et L'action HumaineLcwLchePas encore d'évaluation
- Correction Activité Quantité d'ADNDocument1 pageCorrection Activité Quantité d'ADNWILSON CAMBOURNACPas encore d'évaluation
- De La Generation Spontanee A La TheorieDocument17 pagesDe La Generation Spontanee A La Theoriedimitrio ngoupeyouPas encore d'évaluation
- 135 Biologie J F Beaux G Beaux V BoutinDocument14 pages135 Biologie J F Beaux G Beaux V BoutinGha ZIPas encore d'évaluation
- Document Information General Conservation Des AlimentsDocument28 pagesDocument Information General Conservation Des AlimentsAbdessamad LamnouarPas encore d'évaluation
- Partie 1 M2Document14 pagesPartie 1 M2Amira Amoura100% (1)
- Biologie CellulaireDocument24 pagesBiologie CellulairealmnaouarPas encore d'évaluation
- Ue2 Chapitre 1 - Méthodes en HistologieDocument14 pagesUe2 Chapitre 1 - Méthodes en Histologieakizar ZinoukPas encore d'évaluation
- Extraction Des Acides NucléiquesDocument28 pagesExtraction Des Acides NucléiquesMaï Ssa100% (1)
- Géologie de l'Amérique: Les Grands Articles d'UniversalisD'EverandGéologie de l'Amérique: Les Grands Articles d'UniversalisPas encore d'évaluation
- La vie des abeilles: Prix Nobel de littératureD'EverandLa vie des abeilles: Prix Nobel de littératureÉvaluation : 4 sur 5 étoiles4/5 (41)
- Améliorer votre mémoire: Un Guide pour l'augmentation de la puissance du cerveau, utilisant des techniques et méthodesD'EverandAméliorer votre mémoire: Un Guide pour l'augmentation de la puissance du cerveau, utilisant des techniques et méthodesÉvaluation : 5 sur 5 étoiles5/5 (2)
- Harmonisation Energétique des Lieux: Habitat et haut-lieux sacrés 2020D'EverandHarmonisation Energétique des Lieux: Habitat et haut-lieux sacrés 2020Évaluation : 2.5 sur 5 étoiles2.5/5 (3)
- Encyclopédie de la magie des cristaux, des pierres précieuses et des métauxD'EverandEncyclopédie de la magie des cristaux, des pierres précieuses et des métauxÉvaluation : 4 sur 5 étoiles4/5 (116)
- Le CODE DE DIEU: Le secret de notre passé, la promesse de notre avenirD'EverandLe CODE DE DIEU: Le secret de notre passé, la promesse de notre avenirÉvaluation : 4 sur 5 étoiles4/5 (23)
- Manuel pour les débutants Fabriquez des savons naturelsD'EverandManuel pour les débutants Fabriquez des savons naturelsÉvaluation : 3 sur 5 étoiles3/5 (2)
- Qu'arrive t'il a la terre arable ?: Le problème de l'érosion du sol.D'EverandQu'arrive t'il a la terre arable ?: Le problème de l'érosion du sol.Évaluation : 5 sur 5 étoiles5/5 (1)
- Cahier de jeux de stimulation cognitive: Sujets Alzheimer, désorientés, démences, amnésiesD'EverandCahier de jeux de stimulation cognitive: Sujets Alzheimer, désorientés, démences, amnésiesPas encore d'évaluation
- Atlas permanent de l'Union européenne: 5e édition revue et augmentéeD'EverandAtlas permanent de l'Union européenne: 5e édition revue et augmentéePas encore d'évaluation
- La thérapie libre: La Thérapie de LIBération et Résilience EmotionnelleD'EverandLa thérapie libre: La Thérapie de LIBération et Résilience EmotionnelleÉvaluation : 5 sur 5 étoiles5/5 (1)
- Pourquoi suis-je différent?: Accepter et découvrir la puissance de l'hypersensibilitéD'EverandPourquoi suis-je différent?: Accepter et découvrir la puissance de l'hypersensibilitéPas encore d'évaluation
- Les Graines d'Abricot - Remède de Cancer avec de la Vitamine B17 ?: La Médecine Antique Que l'Industrie Pharmaceutique Moderne CacheD'EverandLes Graines d'Abricot - Remède de Cancer avec de la Vitamine B17 ?: La Médecine Antique Que l'Industrie Pharmaceutique Moderne CachePas encore d'évaluation
- Microbiologie médicale I: agents pathogènes et microbiome humainD'EverandMicrobiologie médicale I: agents pathogènes et microbiome humainÉvaluation : 5 sur 5 étoiles5/5 (1)
- La Conscience Et L'Univers Existent Sans Commencement Ni FinD'EverandLa Conscience Et L'Univers Existent Sans Commencement Ni FinPas encore d'évaluation
- Les 10 Secrets pour une Vie Plus Heureuse avec la Maladie de ParkinsonD'EverandLes 10 Secrets pour une Vie Plus Heureuse avec la Maladie de ParkinsonPas encore d'évaluation
- Traitement des psychonévroses: par la rééducation du contrôle cérébralD'EverandTraitement des psychonévroses: par la rééducation du contrôle cérébralPas encore d'évaluation
- Sédimentologie: Les Grands Articles d'UniversalisD'EverandSédimentologie: Les Grands Articles d'UniversalisPas encore d'évaluation
- Un vent de folie: L'éolien en France : mensonge et arnaque ?D'EverandUn vent de folie: L'éolien en France : mensonge et arnaque ?Pas encore d'évaluation
- Connaître ses réactions émotionnelles, pour maîtriser ses émotionsD'EverandConnaître ses réactions émotionnelles, pour maîtriser ses émotionsPas encore d'évaluation