Vous aimerez peut-être aussi
- +++iKB Hépato-Gastro-Entérologie, Chirurgie Viscérale 6e ÉdDocument607 pages+++iKB Hépato-Gastro-Entérologie, Chirurgie Viscérale 6e ÉdDjallal Hassani100% (3)
- Cours Rénale L2Document72 pagesCours Rénale L2ŚpĕĈíał ØñePas encore d'évaluation
- Histologie de L'appareil-1Document28 pagesHistologie de L'appareil-1qty9jgkpnzPas encore d'évaluation
- 7-Le Foie Et Les Voies BiliairesDocument17 pages7-Le Foie Et Les Voies Biliaireskilo Romeo23Pas encore d'évaluation
- Histologie 2 Partie 4Document110 pagesHistologie 2 Partie 4Salomon JosephPas encore d'évaluation
- Le FoieDocument5 pagesLe FoieLolo KawtchiPas encore d'évaluation
- 7 - Appareil ExcréteurDocument6 pages7 - Appareil ExcréteurClélia RatPas encore d'évaluation
- Les ReinsDocument27 pagesLes ReinsDidi Hmitoche100% (2)
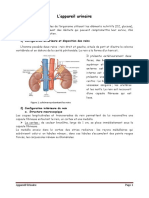
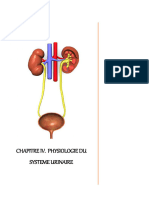
- 7app UrinaireDocument9 pages7app UrinaireDriss AblePas encore d'évaluation
- Histologie Appareil UrinaireDocument48 pagesHistologie Appareil UrinaireKim SeokjinPas encore d'évaluation
- Histologie Des ReinsDocument6 pagesHistologie Des Reinsassil08Pas encore d'évaluation
- U3-H02. Histo Des Reins (15 Pages)Document15 pagesU3-H02. Histo Des Reins (15 Pages)khireddine asmaPas encore d'évaluation
- TP Histo Du ReinDocument14 pagesTP Histo Du Reinchristanklevan2Pas encore d'évaluation
- Physiologie Du ReinDocument15 pagesPhysiologie Du Reinntoutoumelucas17Pas encore d'évaluation
- L'appareil UrinaireDocument65 pagesL'appareil UrinaireAlexandra SparksPas encore d'évaluation
- Commentaires Sur TPDocument3 pagesCommentaires Sur TPMelody's JoolaePas encore d'évaluation
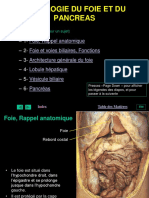
- Foipanc PDFDocument37 pagesFoipanc PDFMohammedAjebliPas encore d'évaluation
- Diaporama Tissus 2Document35 pagesDiaporama Tissus 2Chellouf AminaPas encore d'évaluation
- Histologie Du Tube DigestiveDocument39 pagesHistologie Du Tube DigestivemilimeilysPas encore d'évaluation
- Physio GRD FCT Cours 6 Syst RenalDocument88 pagesPhysio GRD FCT Cours 6 Syst Renalalhassanebah609Pas encore d'évaluation
- 21 - LestomacDocument5 pages21 - Lestomacsameh SakaPas encore d'évaluation
- Le Tube DigestifDocument30 pagesLe Tube Digestifnour nourPas encore d'évaluation
- Ucl Bir TP AnnelidesDocument12 pagesUcl Bir TP AnnelidesWijdene SassiPas encore d'évaluation
- App UrinaireDocument8 pagesApp UrinaireHa NaàPas encore d'évaluation
- Cours Atissulaires 2024Document17 pagesCours Atissulaires 2024Wijdene SassiPas encore d'évaluation
- Cours Du ReinDocument10 pagesCours Du Reinmalakzerizer2005Pas encore d'évaluation
- Physiologie RenaleDocument36 pagesPhysiologie RenaleAnou KalengaPas encore d'évaluation
- Résumé Système UrinaireDocument22 pagesRésumé Système UrinaireMohamed Nassim100% (1)
- Le Système Urinaire SynthèseDocument6 pagesLe Système Urinaire SynthèseSabah MaliatiPas encore d'évaluation
- Anatomie Fonctionelle Rénale 2017 B DioufDocument66 pagesAnatomie Fonctionelle Rénale 2017 B Dioufexauce MondéPas encore d'évaluation
- g2 HIstobashDocument16 pagesg2 HIstobashwjf6kvcm7sPas encore d'évaluation
- Cours 6 HISTOLOGIE DE LAPPAREIL URINAIREDocument9 pagesCours 6 HISTOLOGIE DE LAPPAREIL URINAIREmirvinienPas encore d'évaluation
- Systeme RenalDocument13 pagesSysteme Renala26622058Pas encore d'évaluation
- Systeme Excreteur EtudiantsDocument70 pagesSysteme Excreteur EtudiantsLilà ØūkhPas encore d'évaluation
- Glandes DigestivesDocument28 pagesGlandes Digestivesnour nourPas encore d'évaluation
- Histologie (By Rym)Document40 pagesHistologie (By Rym)rytmPas encore d'évaluation
- 7AS1Document110 pages7AS1zeinebou sidi alhadiPas encore d'évaluation
- Histologie de L'appareil Urinaire1Document9 pagesHistologie de L'appareil Urinaire1Moh LyaliPas encore d'évaluation
- Histo OumaDocument7 pagesHisto OumaAnass VillaPas encore d'évaluation
- Protocole Originale PowerpointDocument59 pagesProtocole Originale Powerpointعمد ابن الفPas encore d'évaluation
- Appareil UrinaireDocument9 pagesAppareil UrinaireTinaPas encore d'évaluation
- Braillon Georges - Le Systeme Nerveux CentralDocument114 pagesBraillon Georges - Le Systeme Nerveux CentralMirela Visan0% (1)
- Cours Glandes Digestives 2014Document76 pagesCours Glandes Digestives 201423azaePas encore d'évaluation
- 5 - Les Capillaires SanguinsDocument3 pages5 - Les Capillaires SanguinsHadjer BmhPas encore d'évaluation
- AppUrinaire HistologieDocument50 pagesAppUrinaire HistologieNIZEYUMUKAMA ProbePas encore d'évaluation
- Physiologie RenaleDocument57 pagesPhysiologie RenaleIr Mukasani BenjaminPas encore d'évaluation
- 1 - Physiologie - Renale +++Document28 pages1 - Physiologie - Renale +++Bilel Dhaouadi100% (1)
- Glandes DigestivesDocument2 pagesGlandes DigestivesAlycia SafaPas encore d'évaluation
- Resume Annelides 1Document2 pagesResume Annelides 1Ny Aina AndriamaroPas encore d'évaluation
- P0612008 PDFDocument101 pagesP0612008 PDFHitomiPas encore d'évaluation
- Histologie de La Rate 2022-2023Document5 pagesHistologie de La Rate 2022-2023Yoü SrãPas encore d'évaluation
- Sémiologie Rénale (Version Provisoire)Document39 pagesSémiologie Rénale (Version Provisoire)Clément PALOUPas encore d'évaluation
- UE Spé 12 Histologie de L'appareil Génital Masculin FellmannDocument21 pagesUE Spé 12 Histologie de L'appareil Génital Masculin Fellmannbilelsta97Pas encore d'évaluation
- 5-Le ColonDocument6 pages5-Le Colonkilo Romeo23Pas encore d'évaluation
- 5-La Paroi GastriqueDocument10 pages5-La Paroi GastriqueSifo GuendouzPas encore d'évaluation
- 2.2la Physiologie RénaleDocument11 pages2.2la Physiologie Rénaleshaiceark.bzhPas encore d'évaluation
- PDF Qe 108 Histo Version FinalDocument14 pagesPDF Qe 108 Histo Version FinalLyna FrikhaPas encore d'évaluation
- Theme 25 Organes Genitaux 3Document41 pagesTheme 25 Organes Genitaux 3Amal NasriPas encore d'évaluation
- Publication Du Cuen Physiologie Et Physiopathologie Renales Chapitre EntierDocument7 pagesPublication Du Cuen Physiologie Et Physiopathologie Renales Chapitre EntierKhady FayePas encore d'évaluation
- Surrénales: Les Grands Articles d'UniversalisD'EverandSurrénales: Les Grands Articles d'UniversalisPas encore d'évaluation
- Appareil circulatoire humain: Les Grands Articles d'UniversalisD'EverandAppareil circulatoire humain: Les Grands Articles d'UniversalisPas encore d'évaluation
- Curs 9 - Sisteme ReproductifDocument16 pagesCurs 9 - Sisteme Reproductifqty9jgkpnzPas encore d'évaluation
- Curs 7 - Les Glandes EndocrinesDocument70 pagesCurs 7 - Les Glandes Endocrinesqty9jgkpnzPas encore d'évaluation
- Histologie de L'appareilDocument33 pagesHistologie de L'appareilqty9jgkpnzPas encore d'évaluation
- Histologie de L Appareil RespiratoireDocument18 pagesHistologie de L Appareil Respiratoireqty9jgkpnzPas encore d'évaluation
- Curs 2 - OchiulDocument39 pagesCurs 2 - Ochiulqty9jgkpnzPas encore d'évaluation
- Curs 4 - Système DigestifDocument42 pagesCurs 4 - Système Digestifqty9jgkpnzPas encore d'évaluation
- Curs 1 - PeauDocument32 pagesCurs 1 - Peauqty9jgkpnzPas encore d'évaluation
- Chapitre2 2Document55 pagesChapitre2 2isabelle gorgeuPas encore d'évaluation
- 2.phase Bucco-OesophagienneDocument10 pages2.phase Bucco-OesophagienneBer MouPas encore d'évaluation
- Proctologie PratiqueDocument122 pagesProctologie Pratiqueproctoaquitaine100% (1)
- Le IléonDocument8 pagesLe Iléonmhzpv82h6gPas encore d'évaluation
- Syndrome de Zollinger Ellison (SZE) : ObjectifsDocument16 pagesSyndrome de Zollinger Ellison (SZE) : Objectifsseed foryouPas encore d'évaluation
- Hépatologie Gastro-Entérologie Et Chirurgie Digestive VG 3ème Edition (Taille Reduite)Document723 pagesHépatologie Gastro-Entérologie Et Chirurgie Digestive VG 3ème Edition (Taille Reduite)Vasilescu IoanPas encore d'évaluation
- Cancer de La VBDocument7 pagesCancer de La VBNizar KaddiouiPas encore d'évaluation
- Postes Choisis Odontologie Semestre de Mai 2020 2Document12 pagesPostes Choisis Odontologie Semestre de Mai 2020 2jzizPas encore d'évaluation
- 2.2 FP Radio Anatomie Du Rectum PDFDocument192 pages2.2 FP Radio Anatomie Du Rectum PDFDjallal HassaniPas encore d'évaluation
- Physiologie Et Exploration Fonctionnelle GastriqueDocument31 pagesPhysiologie Et Exploration Fonctionnelle GastriqueMerou Koubi100% (1)
- Royal MaxFlorDocument2 pagesRoyal MaxFlorstewpitt2Pas encore d'évaluation
- 2 - Abdomen Sans PréparationDocument45 pages2 - Abdomen Sans Préparationayachachoua11Pas encore d'évaluation
- Hémicolectomie Droite Réglée Pour Cancer Par LaparotomieDocument6 pagesHémicolectomie Droite Réglée Pour Cancer Par LaparotomiehoangducnamPas encore d'évaluation
- Cours PROF LATIFA 3APIC S1Document21 pagesCours PROF LATIFA 3APIC S1Youness Boukacem100% (1)
- #Appareil Digestif OK (Histologie Et Embryologie)Document15 pages#Appareil Digestif OK (Histologie Et Embryologie)Moise NdiayePas encore d'évaluation
- Topo Trauma Du FoieDocument55 pagesTopo Trauma Du Foiechergui100% (4)
- 20 Semiologie Des Gros FoiesDocument6 pages20 Semiologie Des Gros FoiesHamda-kati AssowehPas encore d'évaluation
- Tumeur Du Rectum Slides PresentationDocument33 pagesTumeur Du Rectum Slides PresentationKeam PisethsapheareachPas encore d'évaluation
- Ampullome TRTT EndoscopiqueDocument8 pagesAmpullome TRTT EndoscopiqueHouda El MoufidPas encore d'évaluation
- QCM Révision OncologieDocument11 pagesQCM Révision OncologieNabil OuagnouniPas encore d'évaluation
- TP EmbryoDocument10 pagesTP EmbryoJeelian M LukusaPas encore d'évaluation
- Cours, Leçons, Exercices, Cours - Science de La Vie Et de La Terre SVT-LA DIGESTIDocument12 pagesCours, Leçons, Exercices, Cours - Science de La Vie Et de La Terre SVT-LA DIGESTIPape KebePas encore d'évaluation
- Cours Sage Femme 2020 LMDocument71 pagesCours Sage Femme 2020 LMOusmane BARRYPas encore d'évaluation
- Les Cours Des Infirmiers en Toutes Les Modules - ANATOMIE ET PHYSIOLOGIE DE L'APPAREIL DIGESTIFDocument7 pagesLes Cours Des Infirmiers en Toutes Les Modules - ANATOMIE ET PHYSIOLOGIE DE L'APPAREIL DIGESTIFNathan DurandPas encore d'évaluation
- Angiocholite 171007131854Document59 pagesAngiocholite 171007131854Safaa MedPas encore d'évaluation
- Physio Digest L1S2 Ziguinch 2022Document13 pagesPhysio Digest L1S2 Ziguinch 2022antaafall464Pas encore d'évaluation
- Ictères Du Nourrisson Et de L - EnfantDocument4 pagesIctères Du Nourrisson Et de L - Enfantyassine mounajiPas encore d'évaluation
- UntitledDocument2 pagesUntitledYasmina BentorkiPas encore d'évaluation
- Thã©rapie Viscã©rale 2023 2Document208 pagesThã©rapie Viscã©rale 2023 2morgana59Pas encore d'évaluation