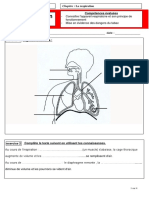
Vous aimerez peut-être aussi
- Physiologie Muscles Striés SquelettiquesDocument16 pagesPhysiologie Muscles Striés SquelettiquessafemindPas encore d'évaluation
- La Physiologie MusculaireDocument51 pagesLa Physiologie MusculaireJoronavalona RASAMIMANANAPas encore d'évaluation
- 2-Physiologie MusculaireDocument44 pages2-Physiologie MusculaireIslęm OuaribPas encore d'évaluation
- Proprietés Mécaniques Du Muscle SquellitiqueDocument9 pagesProprietés Mécaniques Du Muscle SquellitiqueFaculté De Médecine BécharPas encore d'évaluation
- Physiologie CardiaqueDocument55 pagesPhysiologie CardiaqueHadjer Adaida100% (1)
- Anatomie06 PDFDocument3 pagesAnatomie06 PDFHamza AdriPas encore d'évaluation
- Le Cycle CardiaqueDocument8 pagesLe Cycle CardiaqueMohamedBahmed100% (1)
- Muscle Et Physiologie MusculaireDocument20 pagesMuscle Et Physiologie MusculaireBoubker PouperPas encore d'évaluation
- La Contraction MusculaireDocument11 pagesLa Contraction MusculaireJihadi AbbouPas encore d'évaluation
- Liquides Et Electrolytes de L'organismeDocument90 pagesLiquides Et Electrolytes de L'organismeHerby Berthomieux100% (1)
- B04 La VentilationDocument35 pagesB04 La Ventilationfatima kabalanPas encore d'évaluation
- Tissu Musculaire 1Document92 pagesTissu Musculaire 1Adonis Serghini100% (2)
- Tube DigestifDocument72 pagesTube Digestifdrabdelrahim01Pas encore d'évaluation
- Neurophysiologie PDFDocument140 pagesNeurophysiologie PDFPascal Kasero0% (1)
- PCEM1 Physio Cardio VasculaireDocument38 pagesPCEM1 Physio Cardio VasculaireDjallal HassaniPas encore d'évaluation
- Chap1 EucaryoteDocument4 pagesChap1 Eucaryoteslimdamak100% (1)
- Tonus MotriciteDocument15 pagesTonus MotricitecharbelhannaPas encore d'évaluation
- F-Le Système Nerveux VégétatifDocument2 pagesF-Le Système Nerveux Végétatifahlemm100% (1)
- Synapse Et Transmission SynaptiqueDocument10 pagesSynapse Et Transmission SynaptiqueFaculté De Médecine BécharPas encore d'évaluation
- 18 - Amyotrophie Spinales ProgressiveDocument2 pages18 - Amyotrophie Spinales ProgressiveIkram DahmaniPas encore d'évaluation
- 01-Somesthésie 12Document17 pages01-Somesthésie 12nourhoudaPas encore d'évaluation
- Neuroanat-2 0Document83 pagesNeuroanat-2 0seloua kaid100% (1)
- Le SNV (Systeme Nerveux Vegetatif)Document9 pagesLe SNV (Systeme Nerveux Vegetatif)Faculté De Médecine Béchar100% (1)
- Hemorragie - Meningee - LeTresorDesMedecinsDocument4 pagesHemorragie - Meningee - LeTresorDesMedecinsMarMar TsiePas encore d'évaluation
- Fractionné 20220602 1508Document2 pagesFractionné 20220602 1508NounoiPas encore d'évaluation
- 6eme Séance - 2Document42 pages6eme Séance - 2Mohamed AlouanePas encore d'évaluation
- Le Système NerveuxDocument8 pagesLe Système NerveuxazizPas encore d'évaluation
- 3 SomesthésieDocument25 pages3 SomesthésieSahouin Claude MedardPas encore d'évaluation
- Physio CardioDocument16 pagesPhysio CardioWatashi_91Pas encore d'évaluation
- Les Muscles LissesDocument5 pagesLes Muscles LissesHassanElhassanPas encore d'évaluation
- QCM PaDocument1 pageQCM PaBile Djetouan100% (1)
- Biophysique2an Vision2021 PDFDocument18 pagesBiophysique2an Vision2021 PDFMeker NesPas encore d'évaluation
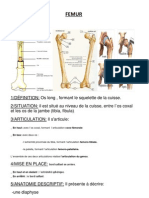
- FemurDocument4 pagesFemurBesma SbPas encore d'évaluation
- FC1 - Les Épithéliums de RevêtementsDocument12 pagesFC1 - Les Épithéliums de RevêtementsModo HecatePas encore d'évaluation
- Cytogenetique pptx-1Document88 pagesCytogenetique pptx-1franck brice badoPas encore d'évaluation
- Evaluation de La Fatigue MusculaireDocument6 pagesEvaluation de La Fatigue MusculairenounouPas encore d'évaluation
- Ok Jeu 5 ELAT S4 Examen Semestriel Cardio-Neuro-Digestif Aout 2011Document2 pagesOk Jeu 5 ELAT S4 Examen Semestriel Cardio-Neuro-Digestif Aout 2011safemind100% (1)
- FC5 Tissu Osseux - Docx 2Document13 pagesFC5 Tissu Osseux - Docx 2Modo HecatePas encore d'évaluation
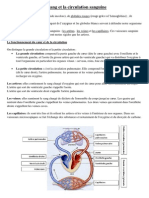
- Le Sang Et La Circulation SanguineDocument1 pageLe Sang Et La Circulation SanguineAgnès FromentPas encore d'évaluation
- Partie I Chap I Le Milieu InterieurDocument15 pagesPartie I Chap I Le Milieu Interieurslimdamak0% (1)
- Td4 Structure de La MP 2019-2020Document2 pagesTd4 Structure de La MP 2019-2020Yugen UniquePas encore d'évaluation
- TSMS - Cours.BH .Chap2. - .I.-.Le - Coeur.et - La.physiologie - CardiaqueDocument7 pagesTSMS - Cours.BH .Chap2. - .I.-.Le - Coeur.et - La.physiologie - Cardiaquepagnapech100% (1)
- Physiologie Du ReinDocument15 pagesPhysiologie Du Reinntoutoumelucas17Pas encore d'évaluation
- Transmission Synaptique PDFDocument8 pagesTransmission Synaptique PDFJean PatatePas encore d'évaluation
- ANATOMIE Appliquée RésuméDocument20 pagesANATOMIE Appliquée RésuméSALMAN KHAYATIPas encore d'évaluation
- 2 Physiologie Du Neurone 1Document45 pages2 Physiologie Du Neurone 1Fofana LancinéPas encore d'évaluation
- Espaces AxillairesDocument3 pagesEspaces AxillairesMoussa Koné100% (1)
- CORRECTION COLLE-JABOT-Homéostasie-01.10.22Document6 pagesCORRECTION COLLE-JABOT-Homéostasie-01.10.22Yann legendrePas encore d'évaluation
- Sclérose en Plaques - NetterDocument9 pagesSclérose en Plaques - NetterS. DonyaPas encore d'évaluation
- S2-Partie I.Chapitre 2.le Systeme MusculaireDocument6 pagesS2-Partie I.Chapitre 2.le Systeme MusculaireZahra ElaoumairiPas encore d'évaluation
- Devoir de Contrôle N°2 - SVT - 3ème Sciences Exp (2010-2011) MR Said Mounir 2Document3 pagesDevoir de Contrôle N°2 - SVT - 3ème Sciences Exp (2010-2011) MR Said Mounir 2bellasfarPas encore d'évaluation
- 2-1les Hormones de La Corticosurrénale Pharmacie PDFDocument100 pages2-1les Hormones de La Corticosurrénale Pharmacie PDFМанель Хадби100% (1)
- NeurophyDocument203 pagesNeurophyEPENGEPas encore d'évaluation
- Le manuel complet sur la scoliose et la chirurgie pour les patients: Un regard impartial en profondeur : qu’attendre avant et pendant l’opération de la scolioseD'EverandLe manuel complet sur la scoliose et la chirurgie pour les patients: Un regard impartial en profondeur : qu’attendre avant et pendant l’opération de la scoliosePas encore d'évaluation
- Le Muscle PDFDocument121 pagesLe Muscle PDFYounes BroukiPas encore d'évaluation
- P Motricité (C. Quairiaux) - 26.03.21Document5 pagesP Motricité (C. Quairiaux) - 26.03.21nasten16fhPas encore d'évaluation
- Physiologie Animale Cours 15Document168 pagesPhysiologie Animale Cours 15g69wwpfw5kPas encore d'évaluation
- Boutonnat Jean p01 PDFDocument61 pagesBoutonnat Jean p01 PDFMoncef PechaPas encore d'évaluation
- MotricitéDocument47 pagesMotricitéMaddy IlPas encore d'évaluation
- Physiologie Du MuscleDocument33 pagesPhysiologie Du MuscleYannick NdriPas encore d'évaluation
- Apports de La Variabilité de La Fréquence Cardiaque Dans L'évaluation de La Charge D'entraînement Et Le Suivi D'athlètes - Aspects Méthodologiques Et Applications PratiquesDocument199 pagesApports de La Variabilité de La Fréquence Cardiaque Dans L'évaluation de La Charge D'entraînement Et Le Suivi D'athlètes - Aspects Méthodologiques Et Applications PratiquesYounes BroukiPas encore d'évaluation
- Chapitre 6 & 7 Dopage Et LutteDocument27 pagesChapitre 6 & 7 Dopage Et LutteYounes BroukiPas encore d'évaluation
- BiomecaniqueDocument35 pagesBiomecaniqueYounes Brouki100% (1)
- Nutrition Cours 4Document20 pagesNutrition Cours 4Younes BroukiPas encore d'évaluation
- Gyn Cologie Du Sport Risques Et B N Fices de L Activit Physique Chez La FemmeDocument512 pagesGyn Cologie Du Sport Risques Et B N Fices de L Activit Physique Chez La FemmeYounes BroukiPas encore d'évaluation
- Anémie Et SportDocument22 pagesAnémie Et SportYounes BroukiPas encore d'évaluation
- Nutrition Cours 1Document19 pagesNutrition Cours 1Younes Brouki100% (4)
- Nutrition Cours 3Document7 pagesNutrition Cours 3Younes Brouki50% (2)
- Nutrition Cours 2Document6 pagesNutrition Cours 2Younes Brouki100% (1)
- Communication SocialeDocument27 pagesCommunication SocialeYounes BroukiPas encore d'évaluation
- Adaptation Cardi-Resp - CoursDocument8 pagesAdaptation Cardi-Resp - CoursYounes BroukiPas encore d'évaluation
- Composantes de Cycle de L'amélioration de La QualitéDocument37 pagesComposantes de Cycle de L'amélioration de La QualitéYounes Brouki100% (1)
- Adaptation Cardo VasculaireDocument48 pagesAdaptation Cardo VasculaireYounes BroukiPas encore d'évaluation
- Axegonadotrope 2019Document54 pagesAxegonadotrope 2019Younes BroukiPas encore d'évaluation
- Cours (2) Vieillissement Et Activité PhysiqueDocument34 pagesCours (2) Vieillissement Et Activité PhysiqueYounes BroukiPas encore d'évaluation
- Cours (1) Physiologie Et Prescription D'exercices Chez La Population FéminineDocument115 pagesCours (1) Physiologie Et Prescription D'exercices Chez La Population FéminineYounes BroukiPas encore d'évaluation
- Axe Gonadotrope FemininDocument70 pagesAxe Gonadotrope FemininYounes BroukiPas encore d'évaluation
- Filière de Soins Spécifiques Des Maladies Non Transmissibles PDFDocument49 pagesFilière de Soins Spécifiques Des Maladies Non Transmissibles PDFYounes BroukiPas encore d'évaluation
- Quels Sport Pour Un AsthmatiqueDocument2 pagesQuels Sport Pour Un AsthmatiqueYounes BroukiPas encore d'évaluation
- CMB Finalisé PDFDocument52 pagesCMB Finalisé PDFYounes BroukiPas encore d'évaluation
- Adaptation Physiologique en Microgravité - Notes de CoursDocument33 pagesAdaptation Physiologique en Microgravité - Notes de CoursYounes BroukiPas encore d'évaluation
- L'Entraînement Physique NotesDocument112 pagesL'Entraînement Physique NotesYounes Brouki100% (2)
- Le Muscle PDFDocument121 pagesLe Muscle PDFYounes BroukiPas encore d'évaluation
- Contrat D'engagement de Joueur Professionnel VFinale 15-16Document8 pagesContrat D'engagement de Joueur Professionnel VFinale 15-16Younes Brouki100% (1)
- Amo - Cours - 2017 CMB PDFDocument82 pagesAmo - Cours - 2017 CMB PDFYounes BroukiPas encore d'évaluation
- PR Bouziane - Anatomie de La Râte - CM PDFDocument33 pagesPR Bouziane - Anatomie de La Râte - CM PDFmomoPas encore d'évaluation
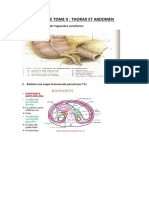
- Splanchnologie Tome IIDocument16 pagesSplanchnologie Tome IINiclette BiayaPas encore d'évaluation
- Anglais 1629072706Document35 pagesAnglais 1629072706Boűmřãh FōüĀdPas encore d'évaluation
- L'appareil UrinaireDocument25 pagesL'appareil UrinairesambaPas encore d'évaluation
- Devoir-De-Classe - SVT 3Document4 pagesDevoir-De-Classe - SVT 3kkkPas encore d'évaluation
- QCM NeuroDocument15 pagesQCM NeuroJonathan JohnsonPas encore d'évaluation
- Exploration de La CorticosurrénaleDocument58 pagesExploration de La CorticosurrénaleHicham BenaoumPas encore d'évaluation
- Resume Cours Cycles Ovarien UtérinDocument2 pagesResume Cours Cycles Ovarien Utérintheeo bebouPas encore d'évaluation
- Recommandation Tumeur SurrenalienneDocument30 pagesRecommandation Tumeur Surrenalienneclaudia nkamPas encore d'évaluation
- DDM OdtDocument10 pagesDDM OdtazouzPas encore d'évaluation
- Laboratoire D'Analyses Medicales Aureslab: Dr. BOUSSAHA Asma Epse Bouguelaa ةحاسوب.أ: ةروتكدلا ةعلقوب ةجوزDocument2 pagesLaboratoire D'Analyses Medicales Aureslab: Dr. BOUSSAHA Asma Epse Bouguelaa ةحاسوب.أ: ةروتكدلا ةعلقوب ةجوزYas MinPas encore d'évaluation
- EMC Anomalies Des Dents TemporairesDocument8 pagesEMC Anomalies Des Dents Temporairestahari nabilaPas encore d'évaluation
- S NaamouneDocument41 pagesS NaamouneMohamed Wassim M'henni0% (1)
- Semiologie UrologiqueDocument5 pagesSemiologie UrologiqueOUMAIMA EL CHAALPas encore d'évaluation
- Examens Histologie (Utile de 48 À La Fin)Document125 pagesExamens Histologie (Utile de 48 À La Fin)heloiserassePas encore d'évaluation
- Muscles de MeridiensDocument13 pagesMuscles de MeridiensJean-Paul RovinotPas encore d'évaluation
- Agencements Intra Et Inter Arcades en Denture PermanenteDocument7 pagesAgencements Intra Et Inter Arcades en Denture Permanenterola bouamaraPas encore d'évaluation
- Histologie de La Rate 2022-2023Document5 pagesHistologie de La Rate 2022-2023Yoü SrãPas encore d'évaluation
- Anatomie HumaineDocument88 pagesAnatomie HumaineAndreea OlimpiaPas encore d'évaluation
- 4a PointsSpecifiquesDocument17 pages4a PointsSpecifiquespassagerx007Pas encore d'évaluation
- 3 - Hemodynamique IntracardiaqueDocument9 pages3 - Hemodynamique IntracardiaqueSerigne Sohibou GayePas encore d'évaluation
- Evaluation de la sphère oro-faciale chez l'enfant âgé de 5 à 6 ans : Elaboration et validation d'un outil d'évaluation auprès de 164 enfants : la B.E.S.O.F - Eclairage sur l'existence d'un lien entre l'oralité verbale et l'oralité alimentaire - Sophie VANNIERDocument203 pagesEvaluation de la sphère oro-faciale chez l'enfant âgé de 5 à 6 ans : Elaboration et validation d'un outil d'évaluation auprès de 164 enfants : la B.E.S.O.F - Eclairage sur l'existence d'un lien entre l'oralité verbale et l'oralité alimentaire - Sophie VANNIERSophie VannierPas encore d'évaluation
- Histologie de L'appareil Urinaire1Document9 pagesHistologie de L'appareil Urinaire1Moh LyaliPas encore d'évaluation
- Immuno31 Lymph BDocument34 pagesImmuno31 Lymph Bzakimoh113BBPas encore d'évaluation
- Devoir de Synthèse N°3 (Corrigé) - Sciences Naturelles - 4ème SC Exp (2008-2009) Mme Raja HabliDocument10 pagesDevoir de Synthèse N°3 (Corrigé) - Sciences Naturelles - 4ème SC Exp (2008-2009) Mme Raja HabliMahdi AchbarPas encore d'évaluation
- TD ReinDocument10 pagesTD ReinDieuboue ArthurPas encore d'évaluation
- T3A2 Acti2 FCTT OvairesDocument3 pagesT3A2 Acti2 FCTT Ovairesbianca fPas encore d'évaluation
- Système CirculatoireDocument96 pagesSystème CirculatoireAnonymous MKSfyYyODP100% (1)
- L'hemogramme Et Son InterpretationDocument74 pagesL'hemogramme Et Son InterpretationOrdi LebonheurePas encore d'évaluation
- EmbryologieDocument4 pagesEmbryologieWissem La TlemcenienePas encore d'évaluation